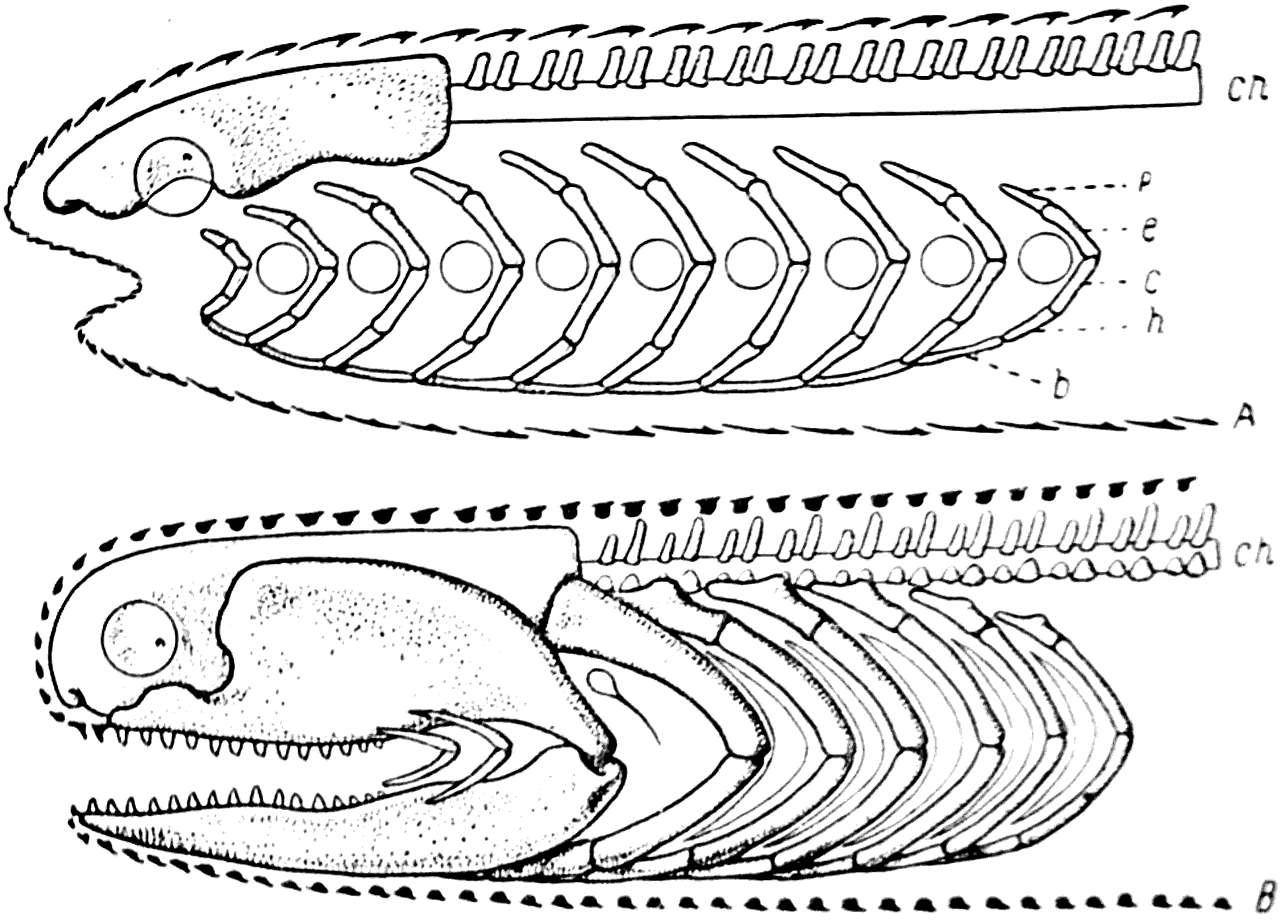
Рис. 40. Схема строения головы бесчелюстного (A) и челюстного (В) позвоночного.
b — basibranchiale; с — ceratobranchiale; ch — chorda; e — epibranchiale; h — hypobranchiale; p — pharyngobranchiale.
Отсутствие в ордовикских отложениях хорошо сохранившихся остатков древнейших представителей Agnatha не дает нам возможности выяснить на палеонтологическом материале все те изменения, которые имели место при превращении бесчелюстных позвоночных в позвоночных, снабженных челюстями.
Однако это обстоятельство не лишает нас права считать, что позвоночные, имеющие челюсти, произошли от каких-то бесчелюстных форм, и пытаться теоретически представить себе все те изменения, которые совершились в скелете позвоночных при переходе от Agnatha к Gnathostomata.
Внутренний скелет первых Gnathostomata, несомненно, был, как и у их бесчелюстных предков, хрящевым, но в нем произошли следующие очень важные изменения.
В позвоночном столбе, по всей вероятности, уже появились хрящевые элементы тел позвонков. Они располагались на нижней стороне хорды в количестве двух пар на каждый позвонок. Таким образом, в состав одного позвонка у Gnathostomata входили две пары дуг и две пары тел. Так построены позвонки у эмбрионов современных акул и это дает нам право предполагать, что именно такое строение имели хрящевые позвонки и у примитивных Gnathostomata (рис. 40).
Рис. 40. Схема строения головы бесчелюстного (A) и челюстного (В) позвоночного.
b — basibranchiale; с — ceratobranchiale; ch — chorda; e — epibranchiale; h — hypobranchiale; p — pharyngobranchiale.
Одновременно с этими усложнениями в позвоночном столбе происходила и очень сильная перестройка хрящевого скелета жаберного аппарата. По существу именно эта перестройка и превратила Agnatna в Gnathostomata.
В процессе формирования хрящевого черепа Gnathostomata первая и вторая жаберные дуги Agnatha подверглись сильной редукции; они потеряли свои верхние и нижние элементы (pharyngobranchiale и hypobranchiale), а две сохранившиеся части (epibranchiale и ceratobranchiale) превратились в тонкие хрящи, заложенные в кожной складке угла рта. Таким образом возникли так называемые губные хрящи, еще сохраняющиеся у многих современных акул.
После редукции двух первых висцеральных дуг жаберные отверстия, располагающиеся позади каждой из них, закрылись.
Значительно более сильным изменениям подверглась третья жаберная дуга. Ее pharyngobranchiale и hypobranchiale редуцировались. Что же касается epi- и ceratobranchiale, то они, наоборот, сильно увеличились в своих размерах и превратились в челюстные хрящи; при этом epibranchiale дало так называемое palatoqoadratum, a ceratobranchiale — mandibulare или cartilago Meckelii (меккелев хрящ). Место соединения этих элементов превратилось в челюстной сустав.
Так как слизистая оболочка рта у зародыша образуется в результате вворачивания эктодермы, то, естественно, она оказывается способной образовывать во рту такие же зубы, какие создавала кожа на теле животного. В результате этого покровы, выстилающие полость рта, у примитивных челюстных позвоночных оказались снабженными зубами. Но здесь зубы уже не могли играть прежнюю роль и с момента образования челюстных хрящей (palatoquadratum и mandibulare) приобрели новую функцию. Прикрепившись своими основаниями к хрящам челюстей, зубы стали теперь уже не орудиями пассивной защиты, а органами, помогающими животному добывать пищу. Это обстоятельство имело огромное значение для всей дальнейшей эволюции Gnathostomata. Эти животные впервые получили возможность не только использовать недоступные их предкам виды пищи, но и более активно вести борьбу за свое существование. Однако следует иметь в виду, что зубы только что возникшего челюстного аппарата на первых этапах своего развития еще не давали рыбам возможности во всех случаях легко отражать нападения современных им хищников. Поэтому все тело древнейших рыб было покрыто, как у их бесчелюстных предков, многочисленными кожными зубами, игравшими роль чешуи и выполнявшими защитную функцию.
Только в процессе дальнейшей эволюции рыб их челюстные зубы постепенно приобретали все большее и большее значение, в то время как кожные зубы, наоборот, теряли свою ценность и, как мы увидим, подвергались постепенной редукции.
После превращения третьей висцеральной дуги в челюстной аппарат располагающееся позади нее жаберное отверстие превратилось в так назывываемое «брызгальце» (spiraculum), при помощи которого глотка сохраняла сообщение с внешней средой.
Четвертая дуга жаберного аппарата Agnatha у Gnathostomata подверглась относительно небольшой перестройке. Она потеряла свой верхний элемент (pharyngobranchiale), но сохранила остальные три, превратившись в так называемую подъязычную дугу. Три элемента этой дуги носят следующие названия: epihyale (hyomandibulare, или ерibranchiale), ceratohyale (hyoideum, или ceratobranchiale), hypohyale (hypobranchiale).
Следующие пять жаберных дуг у Gnathostomata вошли в состав их splanchnocranium без каких-либо особых изменений; жаберные отверстия между ними превратились в жаберные щели.
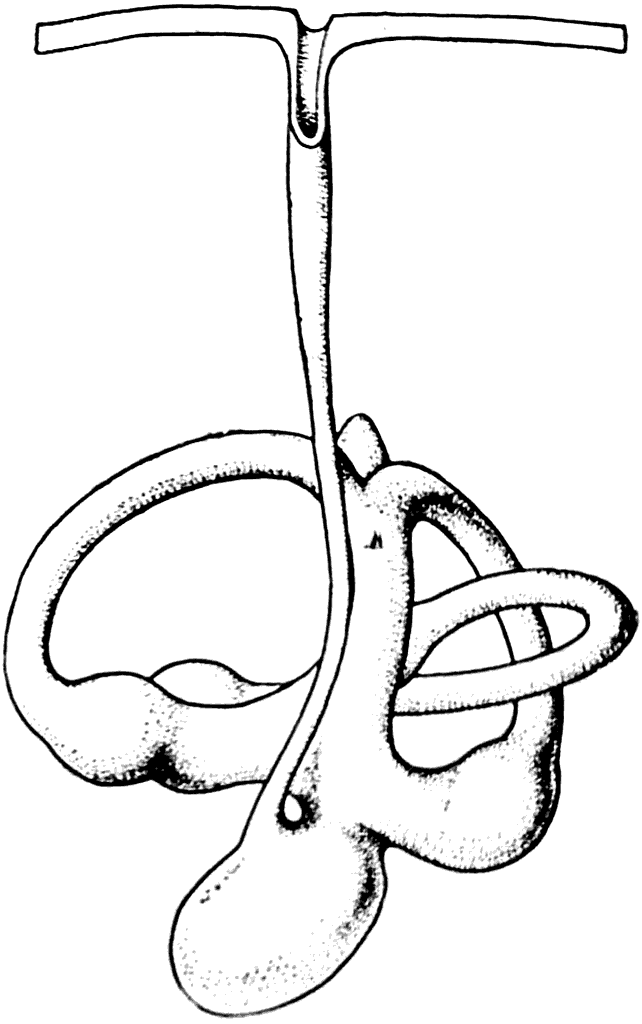
Рис. 41. Перепончатый лабиринт химеры (Chimaera monstrosa).
В слуховом аппарате первых челюстных позвоночных оформились три полукружные канала вместо двух, которые имелись в лабиринте бесчелюстных форм. Такое количество полукружных каналов затем сохранилось совершенно неизменным на протяжении всей долгой эволюции позвоночных, вплоть до человека. Принимая во внимание то обстоятельство, что у некоторых современных примитивных акул полость перепончатого лабиринта при помощи ductus endolymphaticus сообщается с внешней средой (рис. 41), нужно думать, что этой особенностью обладали и примитивные Gnathostomata. Это подтверждается результатами детального изучения некоторых акантодий (Ватсон — D. М. S. Watson, 1937).
Одним из очень важных приобретений древних рыб было образование у них спинных, грудных и брюшных плавников, которые отсутствовали у их предков из класса Agnatha.
Изучение развития плавников у эмбрионов современных рыб показало, что плавники первоначально входили в состав общей кожной складки, которая в виде непарного образования начиналась на спине позади головы, затем окружала хвост и, переходя на нижнюю поверхность туловища, доходила до анального отверстия. Здесь складка разделялась на две части и в виде парного образования тянулась по обе стороны живота до головы животного (рис. 42, A).
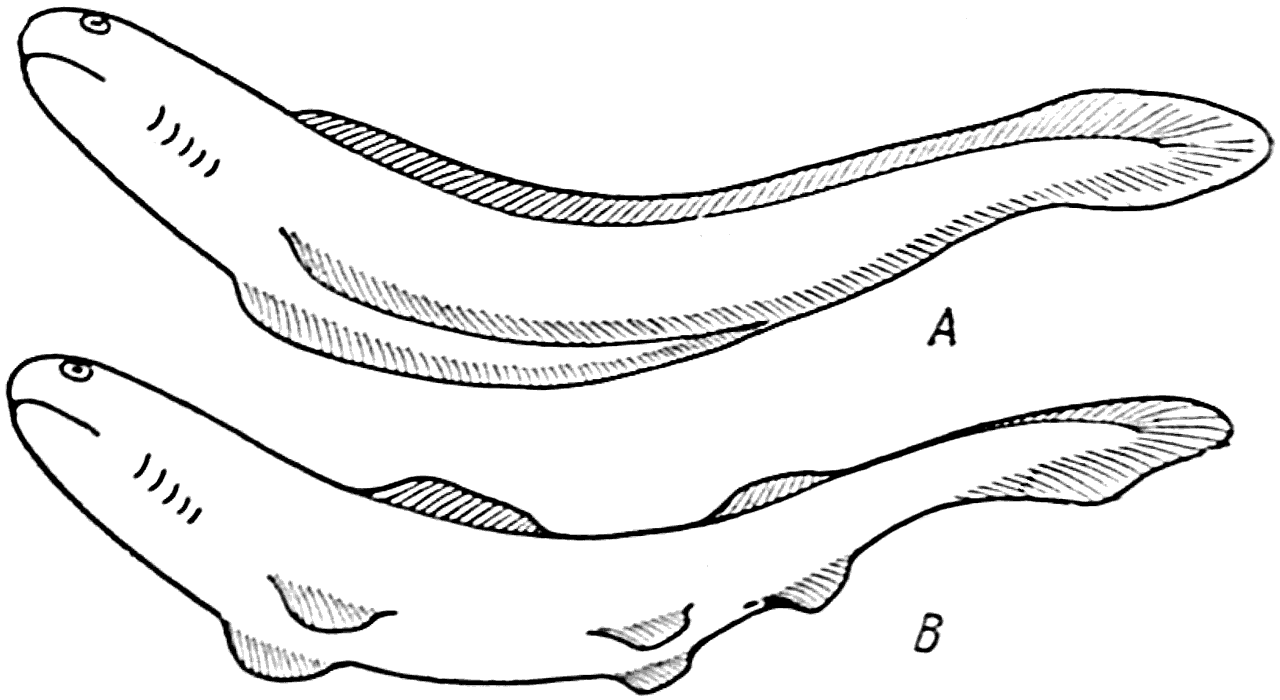
Рис. 42. Образование плавников у рыб (схема).
В дальнейшем исчезновение отдельных участков этой складки привело к выделению двух спинных плавников, хвостового плавника, анального плавника и парных грудных и брюшных плавников (рис. 42, В).
Древнейшими представителями Gnathostomata следует считать акантодий. Это были небольшие рыбы, известные нам еще из силурийских отложений (рис. 17 — Ас).
Все тело силурийских акантодий было покрыто очень маленькими (0,5 мм в длину) плотно прилегающими друг к другу чешуйками, микроскопическое строение которых было очень сложным. Каждая чешуйка по существу состояла из 6—7 генераций зубов, вставленных друг в друга. Первая закладка каждой чешуйки представляла собой очень маленький зуб, состоящий из дентинной основы и покрытый сверху и с боков тонким слоем эмали (рис. 43). .Основание этого первого зуба было сращено с подстилающим его слоем аспидина. На следующем этапе развития чешуи вокруг первого, центрального, зуба возникал новый слой дентина. Он в виде кольца охватывал зуб со всех сторон и наслаивался непосредственно на эмаль, покрывающую его боковые стороны. При этом дентин заходил и на верхнюю поверхность зуба; он откладывался на эмаль и здесь, оставляя свободно только центральную часть ее. Тонкие, снабженные большим количеством ветвей дентинные канальцы во вновь образованном участке дентина обычно перегибались через острый край эмали центрального зуба. Вскоре после образования этого кольцевидного, или, точнее, цилиндрического, слоя дентина на нем возникал тонкий эмалевый колпак. При этом в центральной своей части эмаль ложилась непосредственно на эмаль первого зуба. Одновременно с этим происходило разрастание аспидинового основания чешуи.
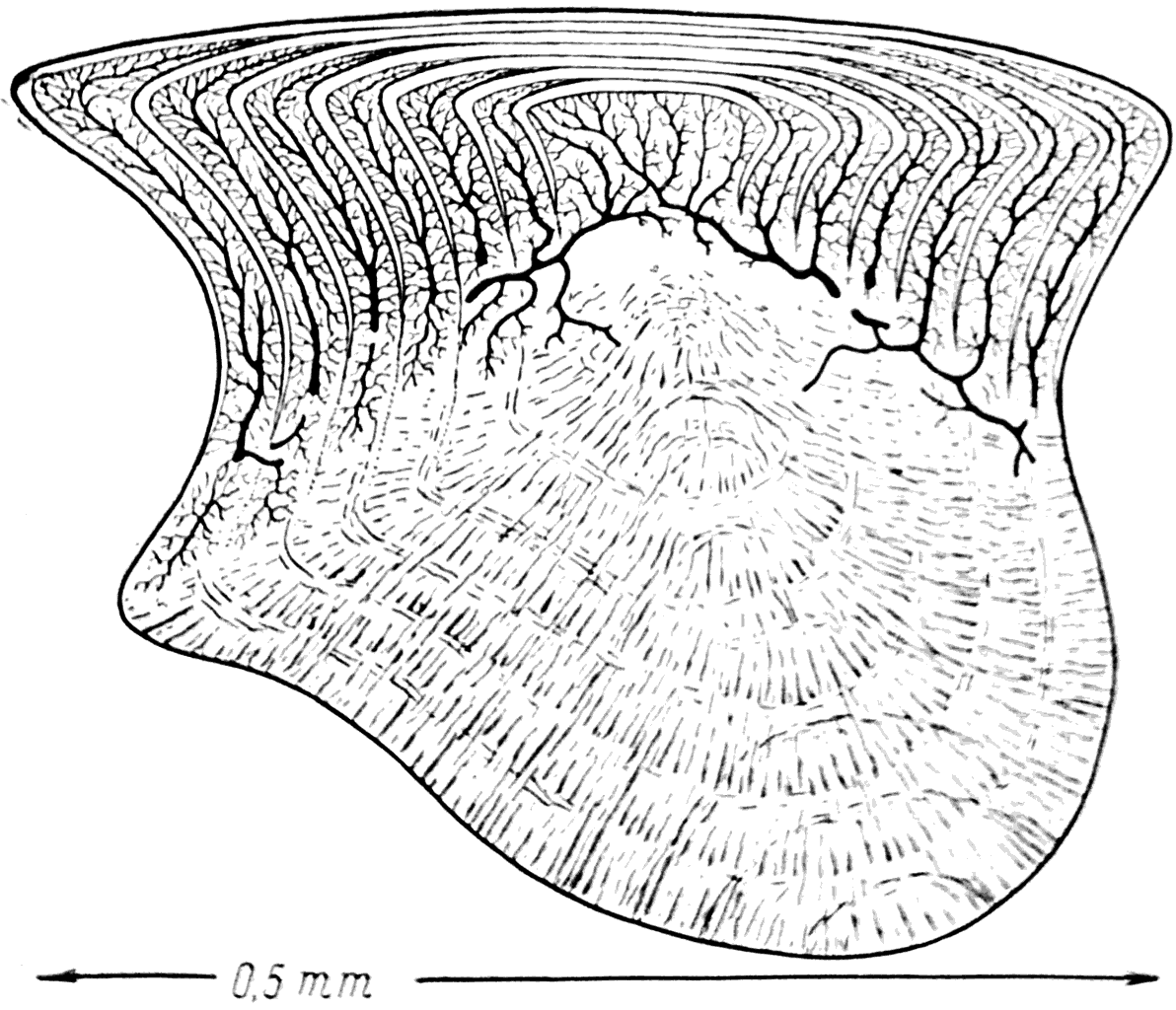
Рис. 43. Вертикальный разрез чешуи Acanthodes.
Последовательный ряд таких наслоений дентина, эмали и аспидина приводил к тому, что чешуя акантодий превращалась в очень сложное образование, состоящее из 6—7 вставленных друг в друга зубов и сращенных со слоистым аспидиновым основанием (рис. 43). Сложность строения чешуи является, разумеется, результатом специализации акантодий, но тот факт, что каждая чешуйка все же представляет собой только видоизмененный зуб (или зубы), одновременно указывает на примитивность этих рыб.
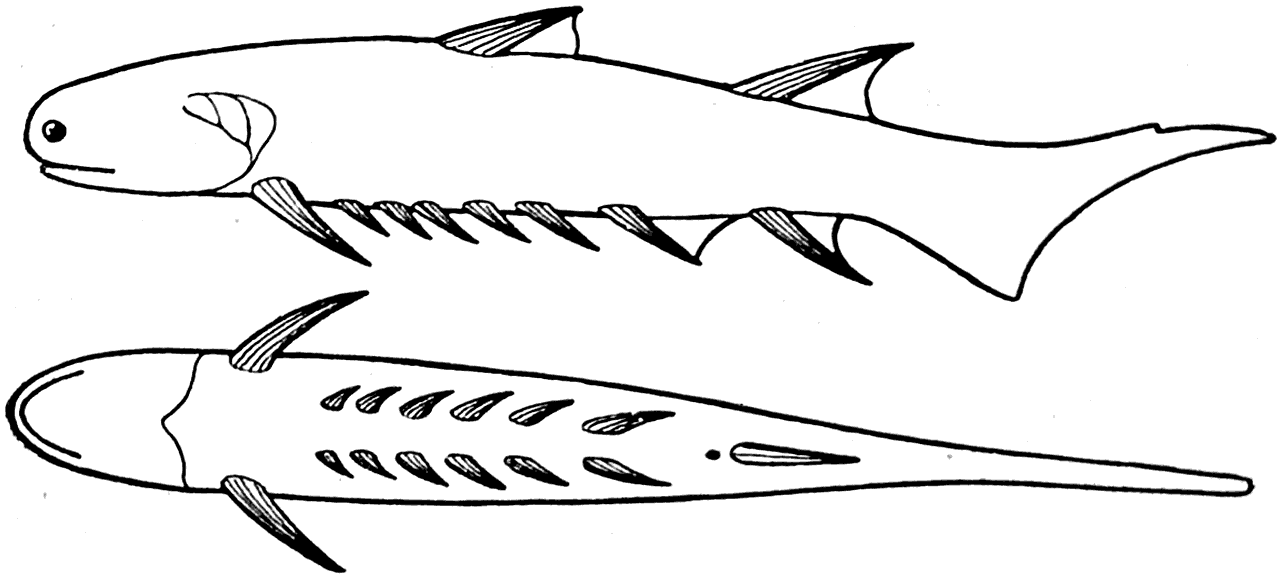
Рис. 44. Euthacanthus macnicoli Powrie. Нижний девон (Ватсон, 1937).
Все плавники у этих древнейших рыб были снабжены острыми шипами. На спине, например, у нижнедевонского Euthacanthus (рис. 44) помещались два шипа, позади которых имелись кожные складки. Такими же складками у этой рыбы были снабжены и анальный и брюшные шипы. Грудные шипы Euthacanthus были лишены кожных складок; не имели кожных складок и пять парных шипов, располагающихся на животе рыбы между ее грудными и брюшными шипами.
Наличие на брюхе акантодий двух рядов шипов, несомненно, указывает на то, что эти шипы развились в парной кожной складке.
Все акантодии были хорошо плавающими рыбами. При этом очень мелкие чешуйки, сидевшие в их коже, не мешали их телу изгибаться при движении. Что же касается острых плавниковых шипов, то они, по-видимому, служили наряду с чешуей неплохими орудиями пассивной защиты от нападений хищников.
Палеонтология в настоящее время не располагает таким материалом, который позволил бы проследить тот эволюционный путь, который прошли Gnathostomata от примитивных форм, сходных с акантодиями, до настоящих костных рыб (Osteichthyes). В силурийских отложениях мы не находим ничего, что давало бы нам возможность выяснить историю развития рыб, снабженных костями.
Для объяснения этого странного явления высказывалось предположение (Ромер — A. S. Romer, 1933), что основные этапы своей эволюции Osteichthyes проделали в верховьях рек, куда в начале девона мигрировали из дельт и озер.
Такую миграцию могли проделать, разумеется, только хорошо плавающие формы, так как жизнь в верховьях рек требовала от рыб способности бороться с быстрым течением. Но это переселение в небольшие притоки рек оказалось очень выгодным для рыб, так как избавляло их от преследований тех врагов, которые, будучи плохими пловцами, не могли проделать такой же миграции (Gigantostraca).
Жизнь в быстро текущих водах верховий рек, по-видимому, была главной причиной все большего усовершенствования плавательных движений, что привело к появлению среди рыб очень сильных пловцов. Кроме того, скоро возникло разделение этих рыб на две группы по способу питания. Одни из них стали питаться в основном растительной пищей, другие превратились в типичных хищников. К сожалению, все этапы этого процесса не могут быть прослежены нами на ископаемом материале, так как никаких остатков силурийских представителей костных рыб мы не находим. Это объясняется тем, что ручьи и небольшие реки, размывающие свои русла, не оставляют осадочных отложений. Поэтому приходится признать, что история развития костных рыб потеряна для нас навсегда. Мы, по-видимому, никогда не сможем найти такие ископаемые остатки, которые позволили бы проследить все интересующие нас фазы развития Osteichthyes. Их кости были перетерты до полного уничтожения в быстро текущих водах верховий рек.
В девоне костные рыбы, уже проделавшие длинную эволюцию в притоках больших рек, неожиданно появились в дельтах и озерах.
Причиной такого возвращения Osteichthyes в места обитания их предков могло быть перенаселение верховий рек. Однако возможно и даже наиболее вероятно, что это было вызвано климатическими особенностями девонского периода. Ряд геологических данных свидетельствует о том, что климат девона был сухим и жарким, по крайней мере в районе нынешних Канады, южной Гренландии, Шпицбергена, Англии, Прибалтики, Воронежской области, Южного Китая, Северной Австралии. На это указывают и некоторые особенности в строении девонских растений (Криштофович, 1941).
Сухой и жаркий климат, естественно, привел к уменьшению сети водных потоков, что вынудило рыб мигрировать в сохранившиеся водные бассейны, т. е. в большие озера и низовья рек.
О сухом климате девона говорят и некоторые особенности самих костных рыб.
Появившиеся в девоне Osteichthyes были частично представлены двоякодышащими (Dipnoi) и кистеперыми (Crossopterygii) рыбами (рис. 17 — D, Cp). И Dipnoi и Crossopterygii, кроме жабр, имели легкие и были снабжены хорошо развитыми грудными и брюшными плавниками, в которых находились наборы хрящевых (или костных) элементов. Эти элементы, к которым прикреплялись мышцы, делали плавники достаточно сильными органами передвижения (рис. 45. A, В).
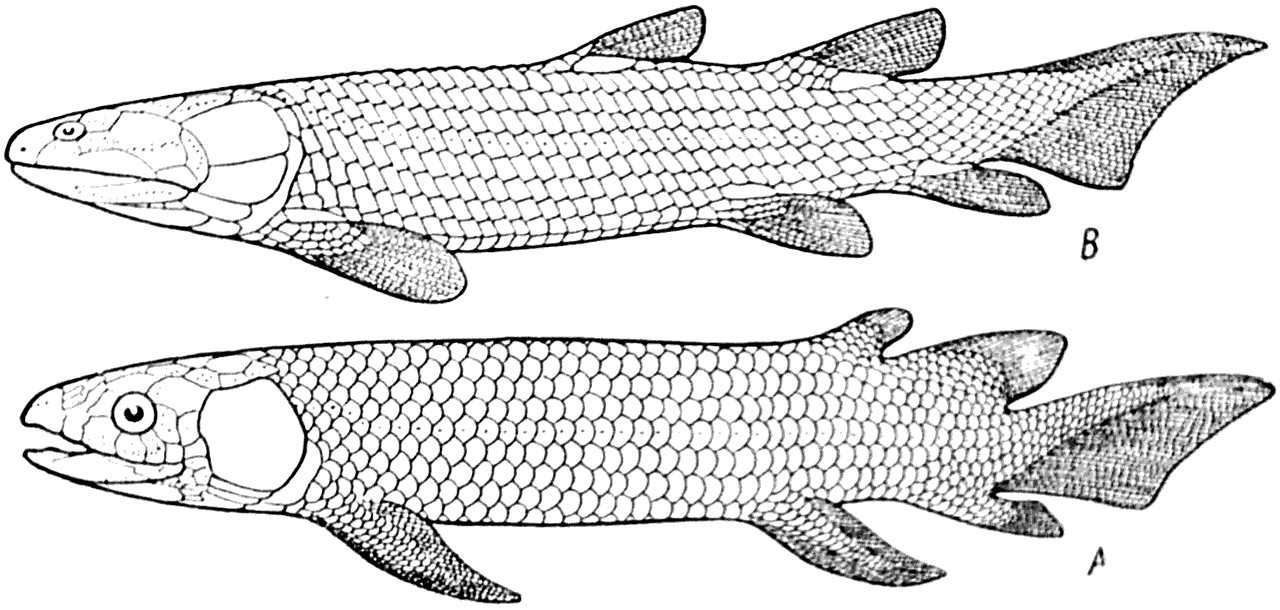
Рис. 45.
А — двоякодышащая рыба Dipterus (Форстер-Купер — Forster-Cooper, 1937); В — кистеперая рыба Osteolepis. Средний девон (Ярвик, 1948).
Легкие двоякодышащих и кистеперых рыб, несомненно, образовались в качестве приспособлений, дающих возможность использовать для газообмена кислород воздуха. Совершенно очевидно, что древнейшие костные рыбы некоторое время находились в таких условиях, в которых одного жаберного дыхания для снабжения организма кислородом было совершенно недостаточно. Такие условия могли создаться только в периодически пересыхающих бассейнах, в воде которых нередко оказывался недостаток кислорода.
Периодическое пересыхание водоемов, по всей вероятности, было причиной появления у двоякодышащих и кистеперых рыб и сильных грудных и брюшных плавников, которые позволяли им в некоторых случаях переползать из пересыхающих водоемов туда, где вода еще сохранялась.
Двоякодышащие и кистеперые рыбы для истории эволюции позвоночных имеют исключительно большое значение, так как изучение их дает возможность выяснить многие вопросы, связанные с проблемой происхождения амфибий.
Dipterus — среднедевонский представитель Dipnoi — был небольшой рыбой; его длина колебалась от 30 до 35 см. В строении этой интересной формы были смешаны как признаки примитивности, так и признаки высокой специализации.
Все тело Dipterus было покрыто циклоидной чешуей, что указывает на некоторую прогрессивность этой рыбы по сравнению с древнейшими формами кроссоптеригий.
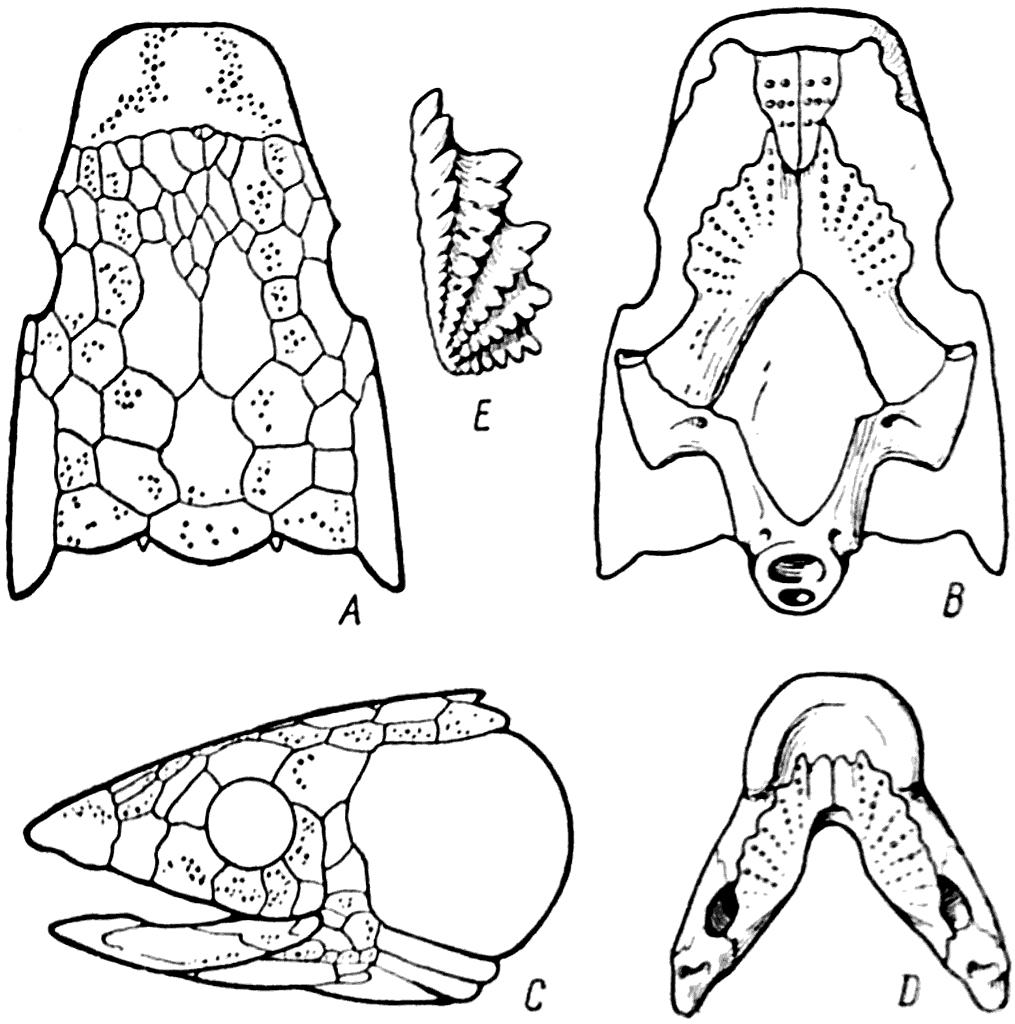
Рис. 46.
А, В, С — череп Dipterus; D — нижняя челюсть Dipterus (Грэхем-Смит и Вестол — Graham-Smith a. Westoll. 1937); Е — зуб двоякодышащей рыбы верхнего девона.
Dipterus имел два спинных плавника и гетероцеркальный хвост. Его грудные и брюшные плавники были очень длинными и в средней своей части были покрыты несколькими рядами чешуй (рис. 45. А). В хрящевом черепе подвергалась окостенению только задняя часть neurocranium (рис. 46, В). Покровные кости, лежащие на chondrocranium, были очень многочисленными, а в переднем отделе черепа срастались в один сплошной щит (рис. 46, A). На нёбе имелось только пять костей: два сошника (vomer), два птеригоида (pterygoideum) и непарный парасфеноид (parasphenoideum) (рис. 46, В); жаберный аппарат был прикрыт большим operculum. Рrаemaxillare и maxillare у Dipterus подверглись полной редукции, поэтому у него отсутствовали краевые зубы. В составе покровных костей нижней челюсти не было dentale; это привело к тому, что и нижняя челюсть была лишена краевых зубов (рис. 46, D). Каналы сейсмосенсорной системы у Dipterus проходили в самом веществе покровных костей и открывались на их наружную поверхность многочисленными отверстиями (рис. 46, А, С).
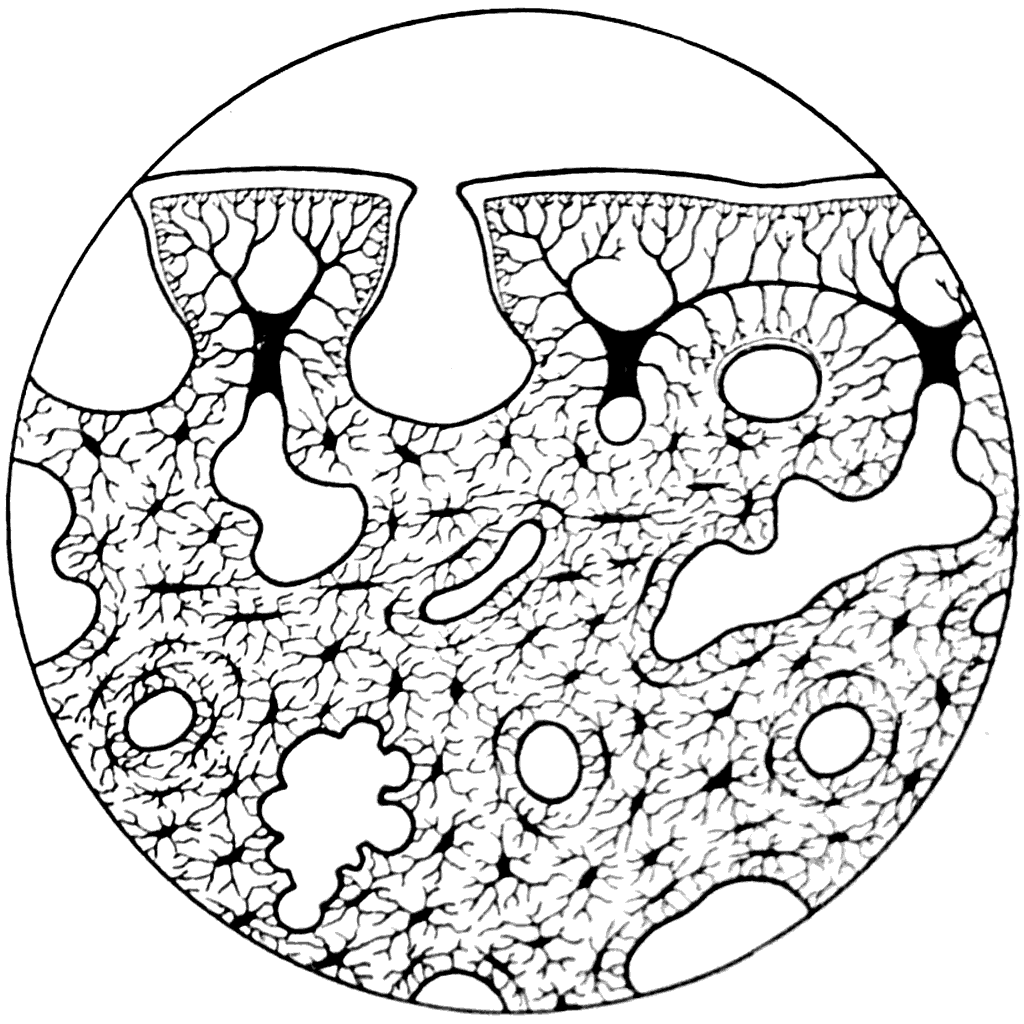
Рис. 47. Вертикальный разрез покровной кости черепа Dipterus (×300) (А. П. Быстров. 1942).
Изучение микроскопического строения чешуи Dipterus и покровных элементов его черепа показало, что эти образования состоят из типичной костной ткани. В ней можно видеть многочисленные полости остеоцитов с тонкими канальцами, отходящими от них в разные стороны (рис. 47). На наружной поверхности как чешуй, так и всех покровных костей Dipterus располагаются очень своеобразные во своей форме кожные зубы. Эти зубы сращены друг с другом и образуют слой, в котором проходят каналы, открывающиеся на поверхность кости круглыми отверстиями. В каждом отдельном зубе имеется небольшая полость пульпы в виде короткого вертикального канала. Стенки зубов толсты и пронизаны дентинными канальцами, ветвящимися на концах. Плоские вершины зубов и их боковые стороны покрыты слоем эмали (А. П. Быстров, 1942).
Среднедевонский Dipterus являлся типичной травоядной рыбой. Необходимость перетирать стебли водных растений требовала какого-то зубного аппарата, и он при отсутствии краевых зубов образовался в результате усиления так называемых шагреневых зубов. Эти очень маленькие конические зубы покрывали у предков Dipterus их птеригоид, лежащий на нёбной поверхности черепа, и praearticulare нижней челюсти. Отдельные шагреневые зубы у Dipterus сильно увеличились в своих размерах и образовали на pterygoideum и praearticulare несколько веерообразно расположенных рядов (рис. 46, В). Между этими рядами у Dipterus еще сохранились очень маленькие шагреневые зубы (рис. 48, А). Дробящие зубы Dipterus отличались большой прочностью (рис. 48, В). Они состояли почти сплошь из дентина и полость их пульпы была представлена системой относительно узких вертикальных каналов, от которых во все стороны расходились дентинные канальцы. Дентин центральной части зуба не содержал дентинных канальцев; здесь протоплазменные отростки одонтобластов исчезли и просветы дентинных канальцев закрылись, в результате чего возник очень прочный витродентин. Наружная поверхность зуба была покрыта слоем эмали.
При закрывании рта ряды дробящих зубов нижней челюсти располагались между рядами соответствующих зубов на нёбной поверхности черепа. Такой своеобразный зубной аппарат, по всей вероятности, позволял Dipterus без труда раздавливать раковины тех маленьких моллюсков и панцыри тех ракообразных, которые вместе с растительностью могли попадать в его рот.
У верхнедевонских представителей двоякодышащих рыб радиально расположенные ряды изолированных конических зубов на нёбе и на нижней челюсти срослись таким образом, что образовали гребнистые зубы, которые являлись еще более мощным аппаратом раздавливания (рис. 46, E).
Dipnoi, часто встречавшиеся в средне- и верхнедевонских отложениях, продолжали существовать, но в меньшем количестве и в карбоне, и в перми. Они жили в пресноводных бассейнах и в течение всего мезозоя. Эти рыбы сохранились и до настоящего времени в Южной Америке (Lepidosiren paradoxa), в Африке (Protopterus annectens) и в Австралии (Neoceratodus forsteri).
Некоторые морфологи конца прошлого столетия (Коп — Соре; Боас — Boas) предполагали, что предков наземных позвоночных (Tetrapoda) следует искать среди двоякодышащих рыб. В настоящее время можно считать установленным, что Dipnoi, уже на первых этапах своей эволюции потерявшие praemaxillare, maxillare и dentale, и приобретшие высокоспециализированный зубной аппарат, предками первых амфибий быть не могли. Этих предков, несомненно, нужно искать только среди кистеперых рыб. Поэтому детальное изучение различных представителей Crossopterygii имеет очень большое значение для решения вопроса о выходе позвоночных из воды на сушу.
Древний и примитивный представитель кистеперых рыб Osteolepis был современником Dipterus. Обе эти рыбы обитали в одних и тех же пресноводных бассейнах среднего девона, на это указывает тот факт, что в соответствующих отложениях их кости нередко бывают перемешаны.
стр. 63