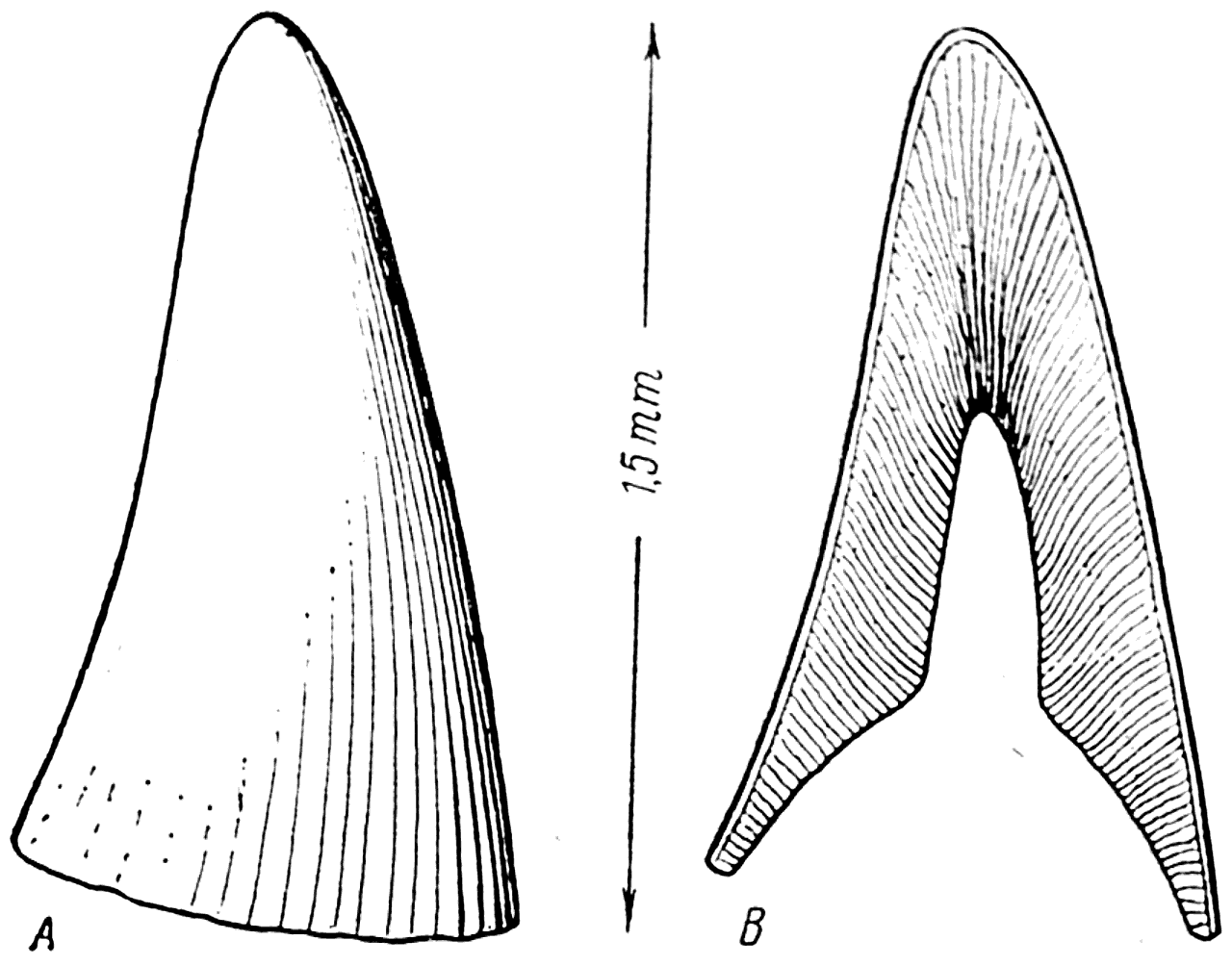
Рис. 18. Кожный зуб Palaeodus (ордовик). А —внешний вид; В — вертикальный разрез (Рогон, 1889).
В отложениях кембрийского периода было найдено немало остатков самых разнообразных беспозвоночных (губки, иглокожие, моллюски, трилобиты), но следов позвоночных обнаружено не было. Это указывает на то, что в это время позвоночные или еще не появились, или их тело еще не имело таких твердых образований, которые могли бы сохраниться в ископаемом состоянии.
Несомненные остатки древнейших позвоночных в настоящее время известны из ордовикских отложений.
В 1889 г. Рогон (Rohon) описал найденные им в глауконитовом песчанике Прибалтики конические зубы, высота которых была равна только 1,5 мм (рис. 18, А). Древность их была около 480 000 000 лет (начало ордовика).
Рис. 18. Кожный зуб Palaeodus (ордовик). А —внешний вид; В — вертикальный разрез (Рогон, 1889).
Изучение микроскопического строения этих зубов показало, что их стенка построена из типичного дентина, пронизанного тонкими дентинными канальцами, а наружная поверхность покрыта слоем эмали; внутри зубов имелась довольно большая полость пульпы (рис. 18, В).
Животное, которому принадлежали эти зубы, получило название Palaeodus и должно быть признано самым древним из известных нам в настоящее время ископаемых позвоночных.
Можно уверенно сказать, что Palaeodus был бесчелюстной формой, т. е. являлся представителем класса Agnatha. Найденные зубы не помещались во рту этого животного, а без сомнения сидели в его коже и играли роль чешуи.
Мы не имеем никакого представления о внешнем виде Palaeodus, так как его тело, кроме кожных зубов, несомненно, не содержало никаких твердых образований, которые могли бы сохраниться в ископаемом состоянии. Что же касается чешуек-зубов, то они оказались разбросанными среди зерен глауконитового песчаника и не давали возможности судить о форме тела животного. Нужно думать, что предки Palaeodus не только во внутреннем скелете, но и на своей коже не имели никаких твердых образований, поэтому у нас по существу нет надежды на то, что мы когда-нибудь найдем их остатки. Таким образом, первые этапы эволюции позвоночных, по всей вероятности, никогда не будут выяснены путем изучения ископаемых форм.
Многочисленные кожные зубы у Palaeodus, несомненно, следует рассматривать как органы защитного значения. Покрывая все тело животного, они в какой-то мере защищали его от врагов. Так как сами бесчелюстные позвоночные не были хищниками и не могли нападать друг на друга, то их преследователей нужно искать среди древних беспозвоночных. Предполагается, что такими врагами могли быть ракоскорпионы (Gigantostraca).
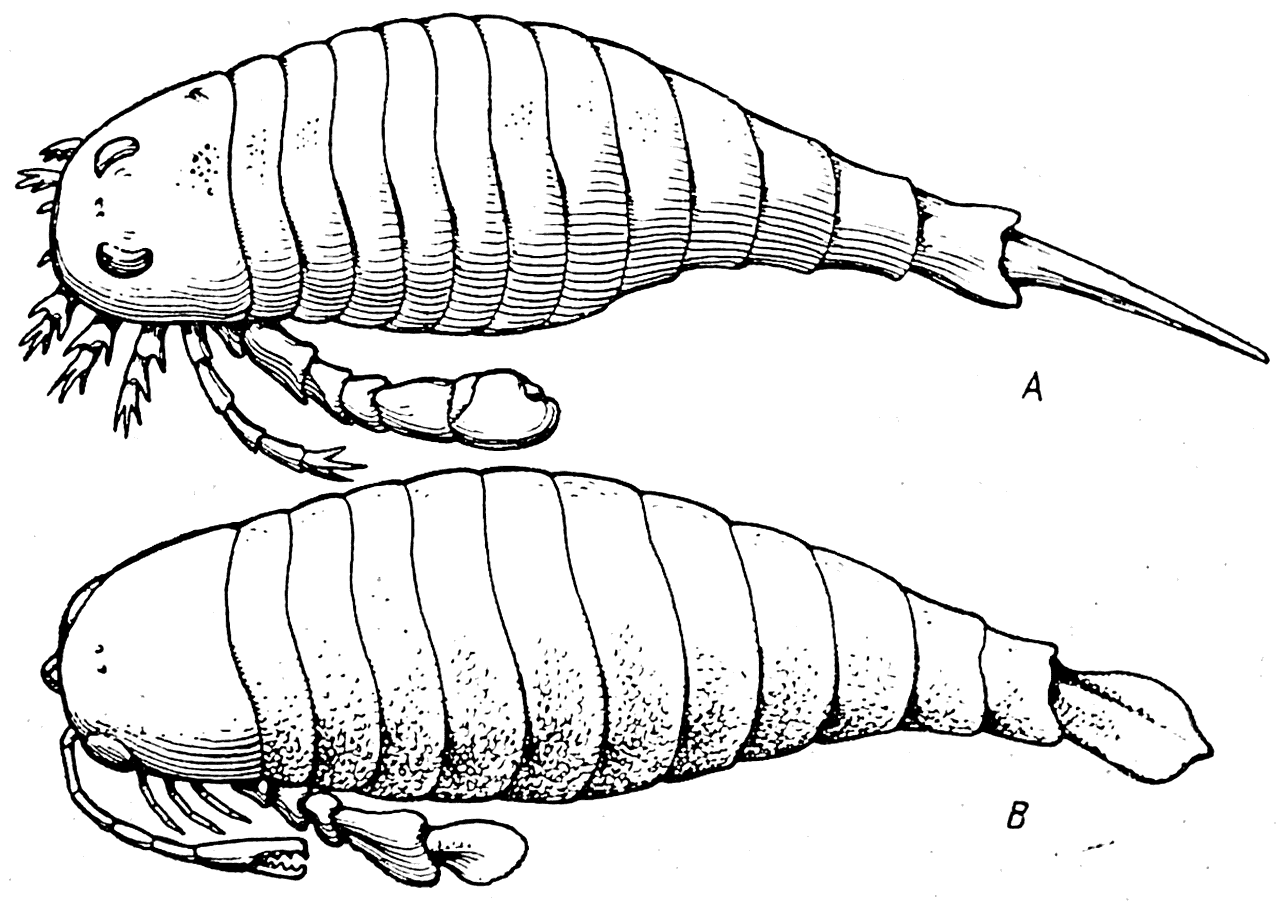
Рис. 19. Силурийские ракоскорпионы. А —Eurypterus fischeri; В — Pterygotus anglicus.
Они появились, по-видимому, еще в кембрии, а в силуре уже в большом количестве населяли водные бассейны (рис. 19). Ракоскорпионы имели членистое тело, покрытое тонким хитиновым панцырем. Последняя пара их конечностей была превращена как бы в весла или лопаты, что давало им возможность медленно плавать и раскапывать ил в поисках своей добычи. У такой формы, как Pterygotus (рис. 19, В), передняя конечность была вооружена клешней, при помощи которой животное могло схватывать свою жертву. Дробящий жевательный аппарат делал этих водных хищников опасными врагами примитивных бесчелюстных позвоночных.
Позвоночные из силурийских отложений известны нам по значительно большему количеству форм, чем позвоночные ордовика.
Подобно современным миногам, все древнейшие формы Agnatha дышали при помощи жабр, которые представляли собой мешки со складчатыми стенками, расположенные по обе стороны глотки. Каждый из этих мешков с одной стороны сообщался с глоткой, с другой — открывался на поверхность кожи. Число жаберных мешков у различных древних форм колебалось от 6 до 15.
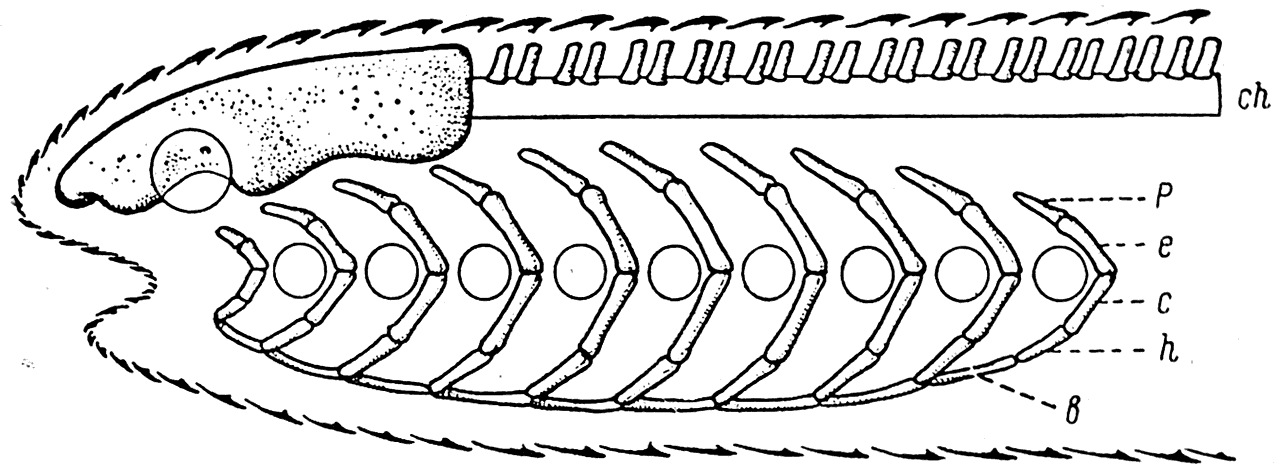
Рис. 20. Схема строения головы Agnatha.
b – basibranchiale; с — ceratobranchiale; ch — chorda; e — epibranchiale; h — hypobranchiale; p — pharyngobranchiale.
Все эти примитивные позвоночные имели несохранявшийся в ископаемом состоянии хрящевой скелет (рис. 20). Их осевой скелет был представлен хорошо развитой хордой (chorda dorsalis). Возможно, что на верхней стороне хорды располагались хрящевые дуги позвонков, каждая из которых состояла из правой и левой половин. Эти половины не были сращены друг с другом. При этом одному позвонку соответствовали, как у современных миног, две такие дуги. Таким образом, каждый позвонок был по существу двойным (диплоспондильным) и состоял из четырех самостоятельных хрящевых элементов. Тела позвонков, по всей вероятности, еще отсутствовали, как они отсутствуют у современных круглоротых (миноги и миксины).
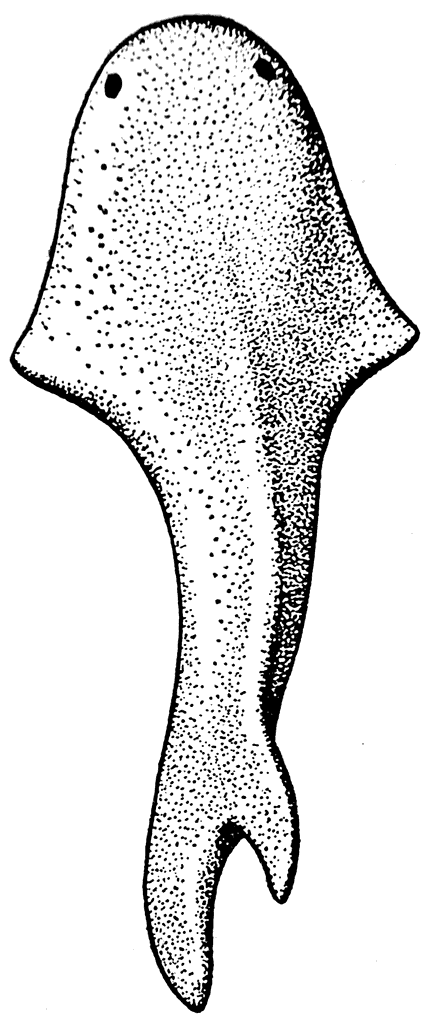
Рис. 21. Thelodus (силур).
Лишенный челюстного аппарата череп (chondrocranium) древнейших позвоночных состоял из хрящевой мозговой коробки (neurocranium) и ряда жаберных дуг, между которыми располагались отверстия жаберных мешков (splanchnocranium). Каждая из жаберных дуг, вероятно, состояла из четырех элементов: pharyngobranchiale, epibranchiale, ceratobranchiale и hypobranchiale. Последний из этих элементов своим нижним концом присоединялся к непарному хрящу, носящему название basibranchiale.
Приспособление бесчелюстных позвоночных к условиям жизни в пресноводных бассейнах уже в силуре привело к тому, что многие из них превратились в придонные формы и по своему внешнему виду стали напоминать скатов. Они рылись в илистом дне водоемов, питаясь продуктами распада растений и жившими в илу мелкими беспозвоночными.
Такой придонной формой был, например, Thelodus (рис. 21). Он имел широкую уплощенную голову, относительно узкое туловище, заканчивающееся гетероцеркальным хвостом. Размеры этого животного были невелики: он имел около 8 см в длину. Его тело было покрыто многочисленными и хорошо развитыми кожными зубами, которые по своей форме сильно отличались от кожных зубов Palaeodus.
Каждый кожный зуб Thelodus имел овальные очертания с заостренным концом, который был направлен назад (рис. 22, A); длинная ось овала равнялась только 0,75 мм. Верхняя поверхность зуба была относительно плоской. Основание зуба, входившее в кожу животного, было сужено (рис. 22, B, С). Как показало изучение шлифов под микроскопом, стенка такого зуба состояла из типичного дентина, содержащего многочисленные дентинные канальцы. Сверху и с боков зуб был покрыт слоем эмали (рис. 22, D).
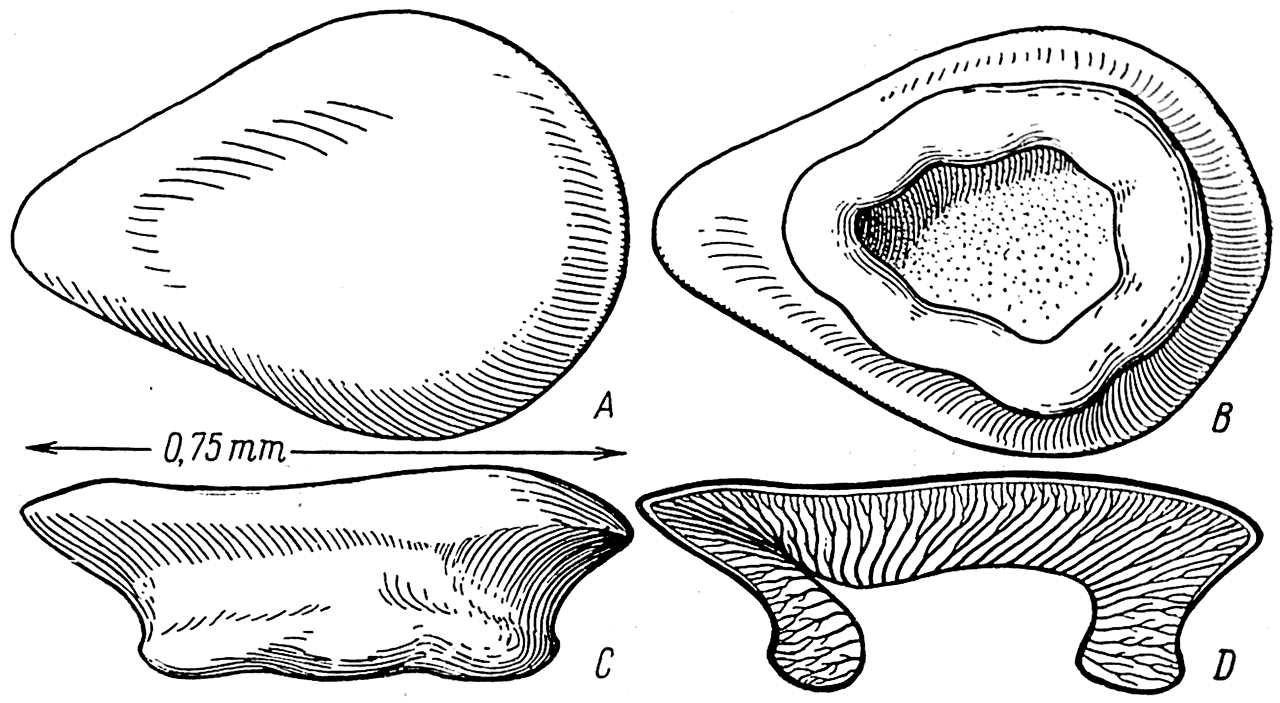
Рис. 22. Чешуя Thelodus. А — вид сверху; В — вид снизу; С — вид сбоку; D — вертикальный разрез (силур) (А. П. Быстров, 1955).
Такие чешуйки-зубы, несомненно, в какой-то мере защищали Thelodus от современных ему хищных форм беспозвоночных. Однако такая защита, по-видимому, не была достаточно надежной, поэтому у некоторых представителей силурийских Agnatha вместо многочисленных кожных зубов возник сплошной прочный панцырь, покрывающий переднюю часть их тела.
Этот панцырь, по всей вероятности, образовался следующим путем. Изучение строения кожных зубов у различных силурийских Agnatha, относящихся, как и Thelodus, к отряду Coelolepida (рис. 17 — C, доказали, что у некоторых из них каждый кожный зуб имел на переднем краю своего основания более или менее сильно развитый отросток (рис. 23, А). Этот отросток не состоял из кости (в нем никогда не удавалось обнаружить полостей остеоцитов); он не был построен и из дентина (в нем не было дентинных канальцев); но вся его масса была пронизана беспорядочно разбросанными тонкими канальцами, иногда собранными в пучки (рис. 23, В). Судя по расположению этих канальцев и их ширине, можно уверенно сказать, что у живого животного в них помещались коллагеновые волокна соединительной ткани. Эти каналы по существу ничем не отличаются от шарпеевских каналов кости.
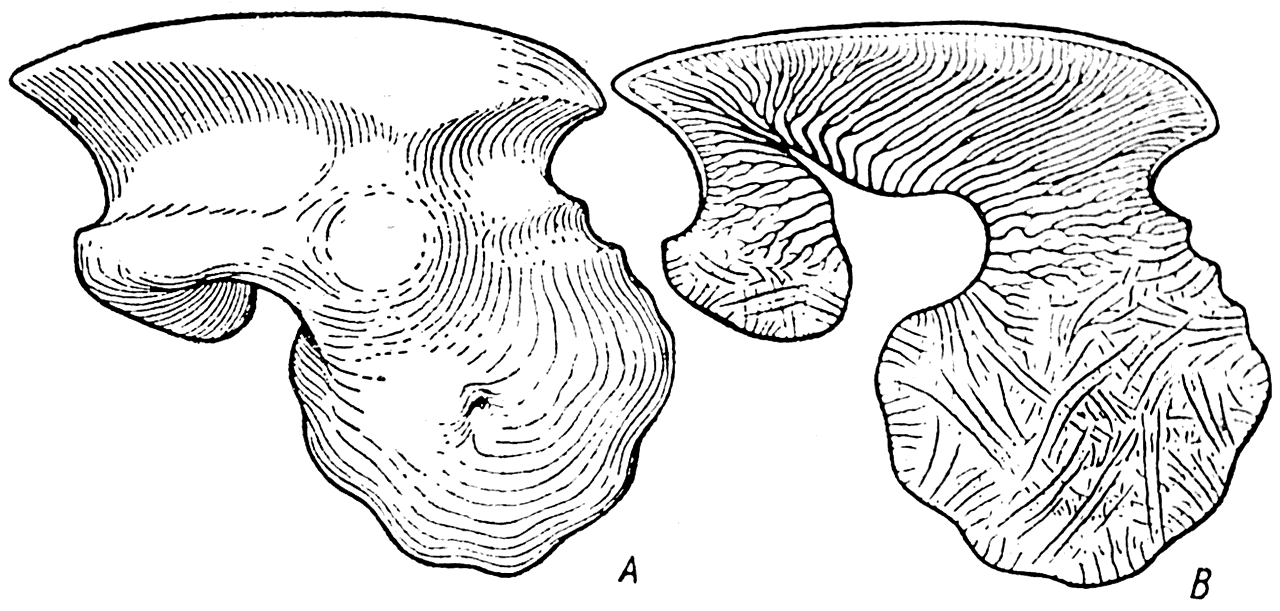
Рис. 23. Чешуя одного из представителей целолепид (Bystrowia aspidinella Berg.). А — вид сбоку; В — вертикальный разрез (силур) (А. П. Быстров, 1955).
Такое твердое вещество — не дентин и не кость, — уже не встречающееся в скелетных образованиях у современных позвоночных, получило название аспидина.
Возможно, что появление аспидиновых наростов на основаниях кожных зубов у некоторых представителей целолепид было первым шагом по пути к образованию сплошного панцыря. Если мы допустим, что затем последовало срастание аспидиновых отростков отдельных зубов друг с другом, то это естественно должно было привести к возникновению на теле животного сплошного панцыря.
Подтверждением этого, мне кажется, может служить головной щит представителя отряда Heterostraci (рис. 17 — H), Tolypelepis undulata Pander.
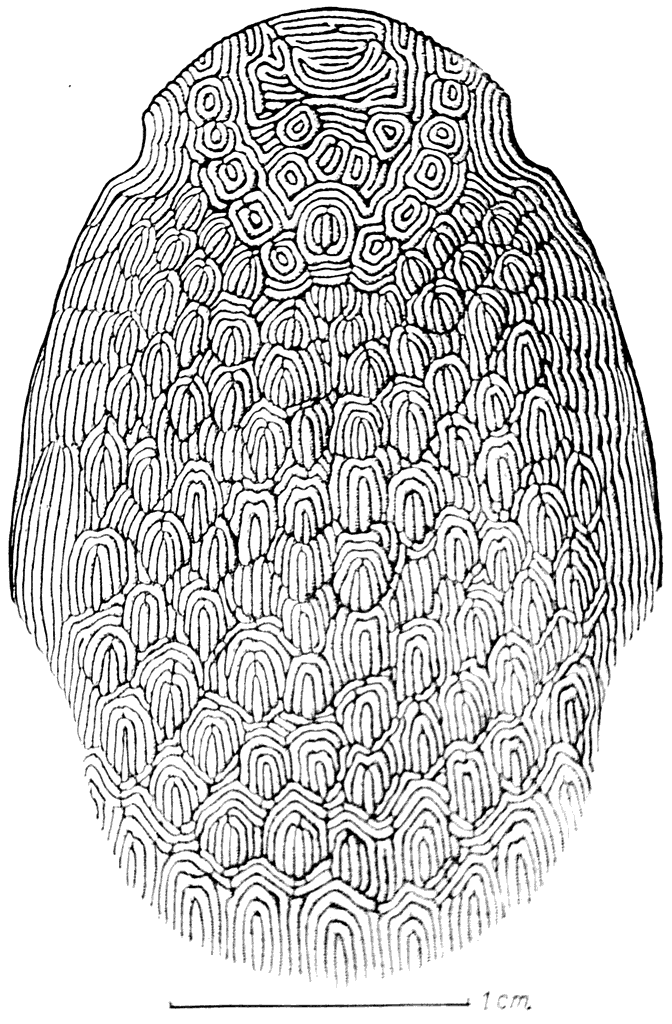
Рис. 24. Щит Tolypelepis undulata (силур).
Tolypelepis был небольшим животным; его головной щит имел немного больше 2 см в ширину. Толщина щита была незначительной, она равнялась только 0,7 мм. Вся наружная поверхность панцыря Tolypelepis была покрыта многочисленными гладкими валиками, собранными в отдельные группы (рис. 24).
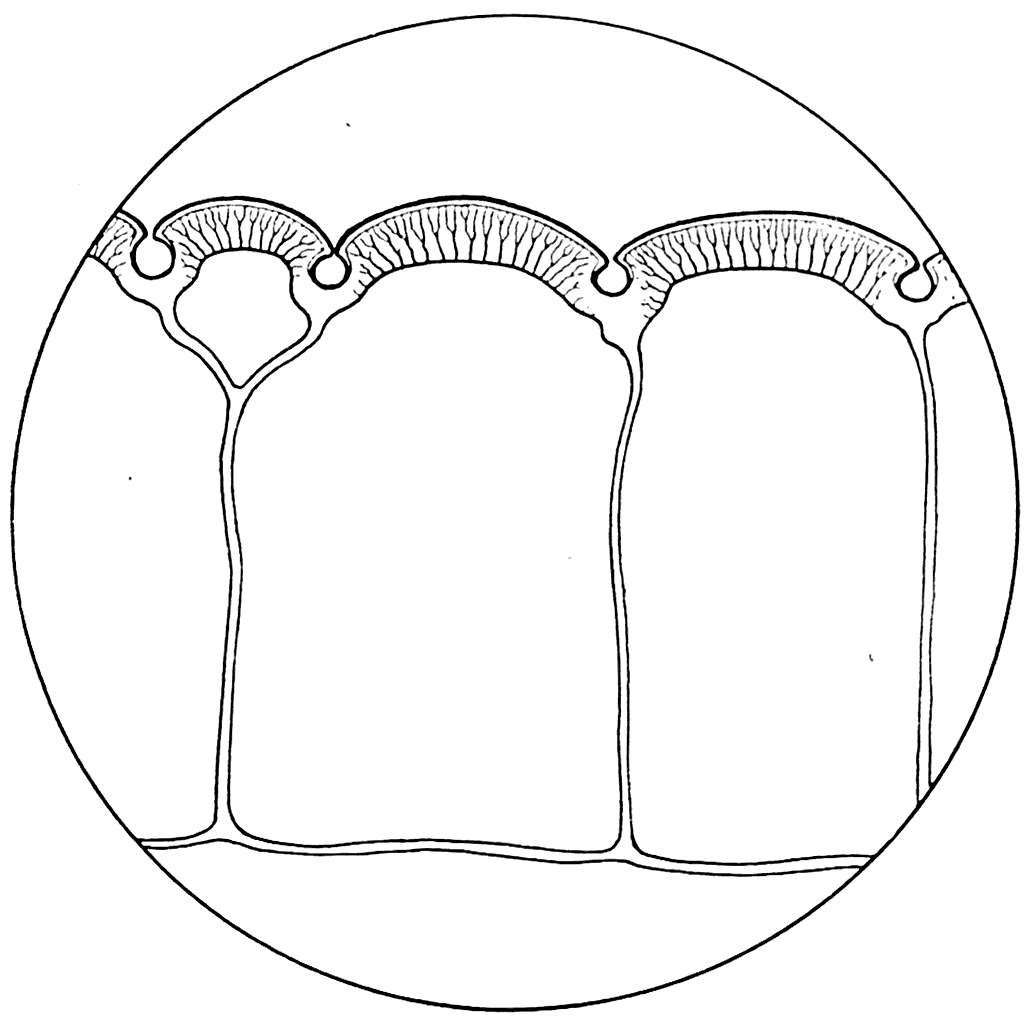
Рис. 25. Вертикальный разрез панцыря Tolypelepis undulata (×300) (А. П. Быстров, 1955).
Изучение микроскопического строения этого панцыря позволило установить, что он состоит из сильно видоизмененных и сросшихся друг с другом кожных зубов (рис. 25). Оказалось, что каждый валик на панцыре представляет собой изогнутую пластинку дентина, пронизанную дентинными канальцами и покрытую тонким слоем эмали. Места сращений каждой пары таких зубов соединены с тонкими вертикальными перегородками, которые в свою очередь сращены с такой же тонкой базальной пластинкой. И перегородки и базальная пластинка в панцыре Tolypelepis построены из аспидина.
Несмотря на тонкость аспидиновых перегородок и базальной пластинки, которые ограничивали заполненные рыхлой соединительной тканью камеры, головной щит Tolypelepis все же мог лучше защищать животное от нападений хищников, чем изолированные кожные зубы, покрывавшие тело целолепид.
Сходное строение имели панцырные щиты и у таких Heterostraci силура, как Poraspis polaris и Pteraspis rostrata. Обе эти формы, несомненно, находились в родственной связи с Tolypelepis, но были значительно более прогрессивными.
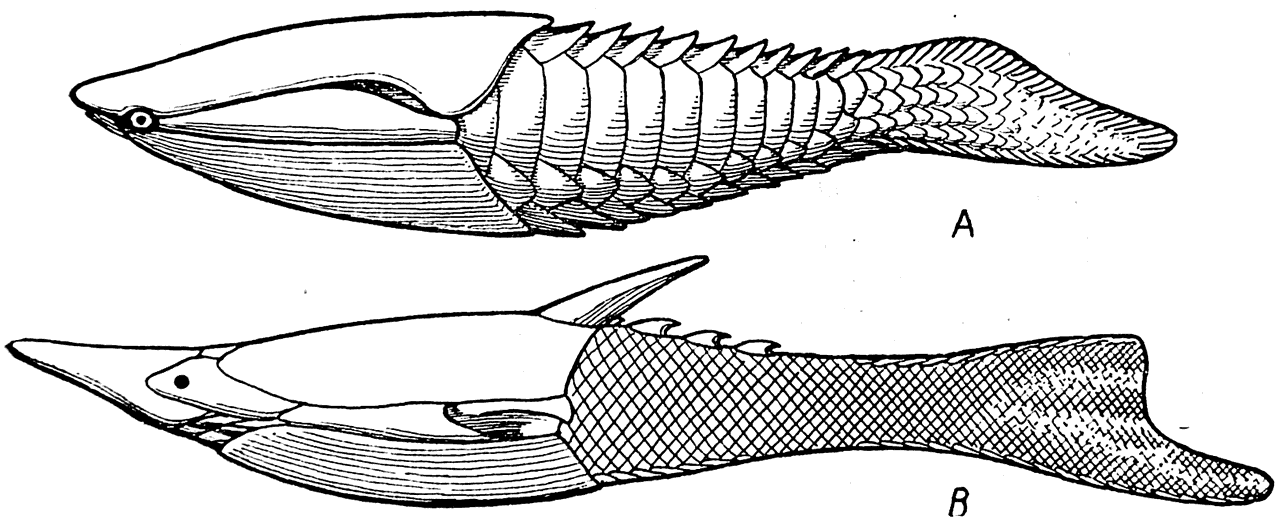
Рис. 26.
А — Poraspis polaris; В — Pteraspis rostrata (силур).
Poraspis (рис. 26, А) достигал 8 см в длину, его голова сверху была прикрыта сплошным щитом, на переднем конце которого справа и слева имелись небольшие вырезки для глаз. На каждой стороне позади глаза располагалась длинная относительно узкая панцырная пластинка, прикрывавшая жаберный аппарат. Жаберным отверстием у Poraspis служила щель между задним концом этой пластинки и головным щитом. Снизу голова животного была защищена большим вентральным щитом. Все тело Poraspis было покрыто большими чешуями; хвост был гипоцеркальным.
Длина Pteraspis (рис. 26, В) колебалась от 13 до 15 см. Это животное обладало большим количеством панцырных элементов на голове, чем Poraspis, и было снабжено острым шипом, сидящим на заднем краю головного щита. Туловище Pteraspis было покрыто мелкими ромбическими чешуями.
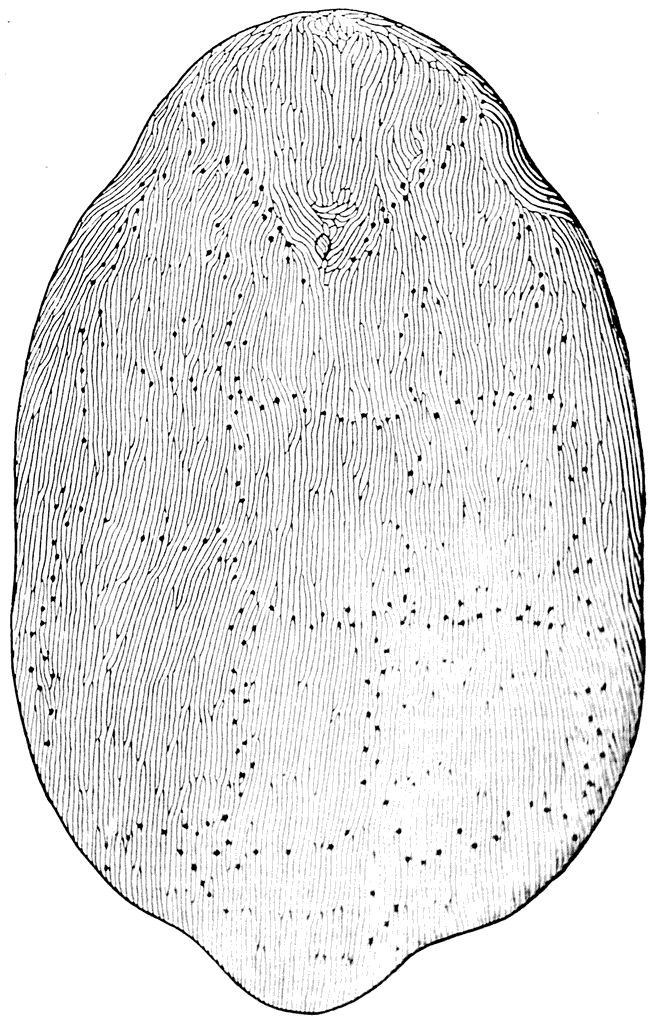
Рис. 27. Щит Poraspis polaris (Киэр и Гейни, 1906).
На наружной поверхности панцырных элементов обеих этих форм можно видеть узкие длинные блестящие валики, ширина которых равняется только 0,2 мм (рис. 27). Изучение вертикальных разрезов панцыря под микроскопом показало, что эти валики построены, как зубы. Они состоят из дентина, пронизанного ветвящимися на концах дентинными канальцами, и покрыты эмалью (рис. 28). Под слоем этих зубов в щитах Poraspis и Pteraspis имеются, как и у Tolypelepis, вертикальные перегородки, присоединяющиеся к базальной пластинке. И перегородки, и базальная пластинка у них несравненно толще, чем у Tolypelepis, поэтому под микроскопом хорошо видно, что эти образования построены из типичного аспидина, в котором легко обнаруживаются многочисленные шарпеевские каналы.
Более прочный панцырь Poraspis и Pteraspis, несомненно, являлся более надежной защитой от нападений хищников, чем панцырь Tolypelepis.
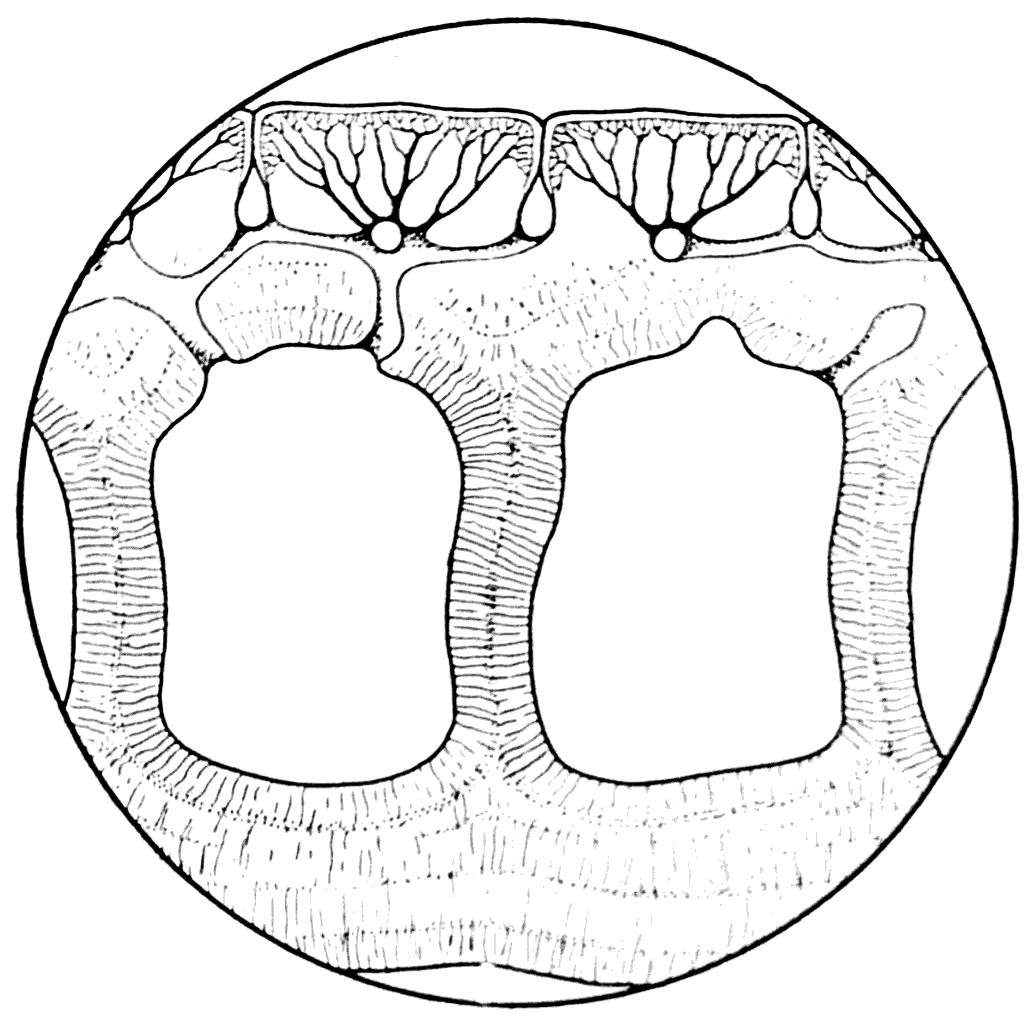
Рис. 28. Вертикальный разрез панцыря Poraspis polaris (×300) (А. П. Быстров, 1955).
У Poraspis и Pteraspis как, вероятно, в у всех представителей силурийских Agnatha, была хорошо развита так называемая сейсмосенсорная система. Она была образована каналами, проходящими в самом веществе панцыря и открывающимися при помощи ряда коротких трубок на наружную поверхность (рис. 29, А). После удаления верхнего слоя щита сейсмосенсорная система Poraspis и Pteraspis обнаруживается в виде сети узких каналов (рис. 29, B), а на поперечных разрезах панцыря ее отдельные участки представляются отверстиями, располагающимися в аспидиновых перегородках под слоем кожных зубов (рис. 29, С).
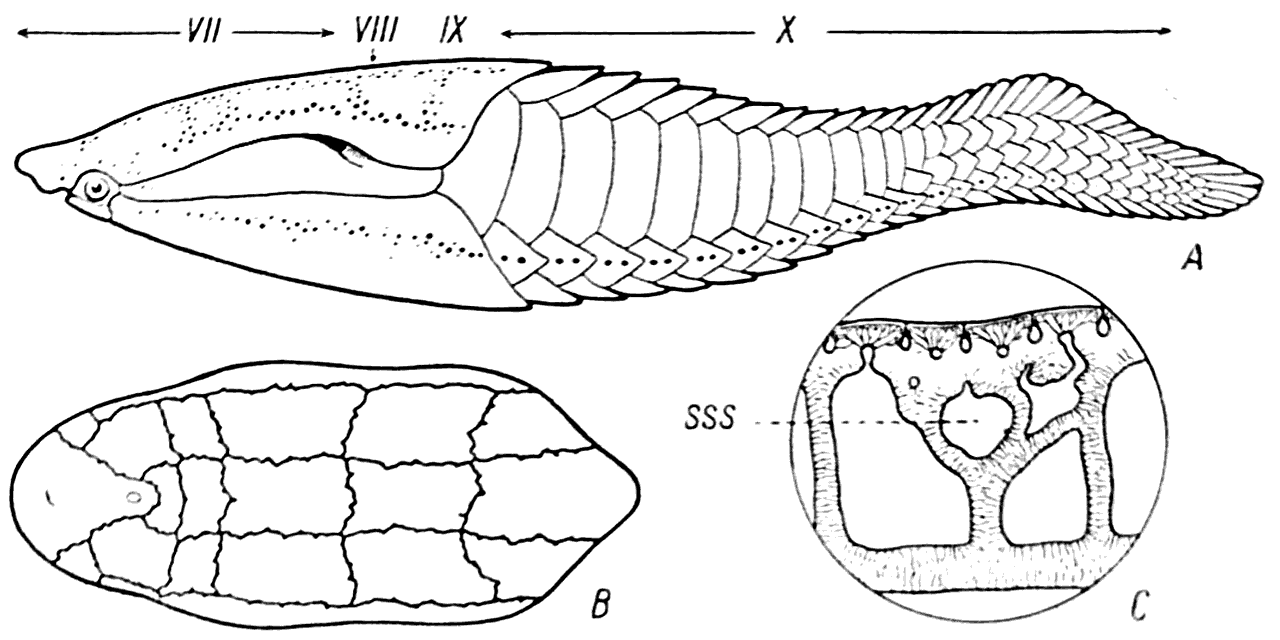
Рис. 29.
А — Poraspis polaris; В — каналы сейсмосенсорной системы в его дорзальном панцырном щите; С — вертикальный разрез щита; SSS — сейсмосенсорный канал.
В этих каналах помещались эпителиальные трубочки, в некоторых участках которых имелись группы особых чувствительных клеток, снабженных волосками.
О структуре этих своеобразных органов мы можем судить, разумеется, только на основании изучения строения таких же образований у современных рыб.
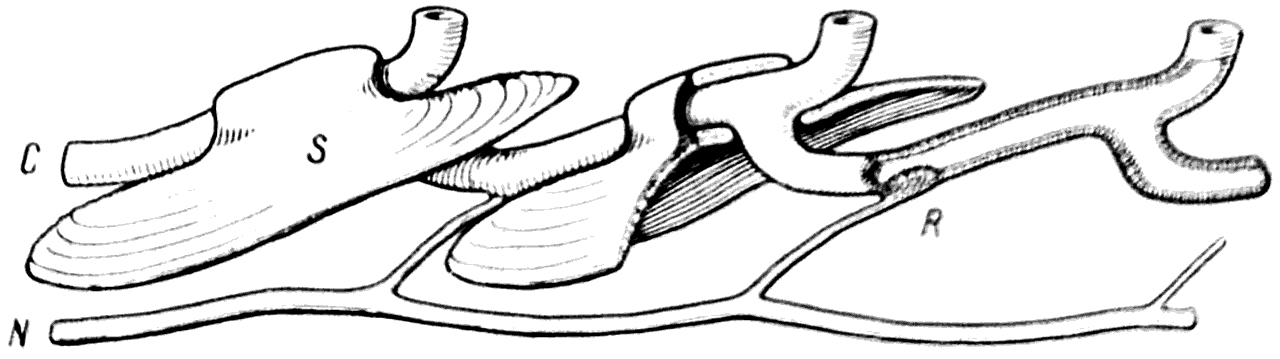
Рис. 30. Схема строения сейсмосенсорной системы у современных рыб.
С — канал; N — нерв; R — рецептор; S — чешуя.
Сейсмосенсорная система рыб образована эпителиальными каналами, заложенными в покровных костях черепа и под чешуей (рис. 30), Эти каналы при помощи коротких ветвей открываются на поверхность кожи. В некоторых участках стенки каналов имеются скопления эпителиальных клеток, снабженных волосками и образующих своеобразные рецепторы.
К каждому такому рецептору направляется нерв и охватывает своими ветвями его чувствительные клетки (рис. 31, A).
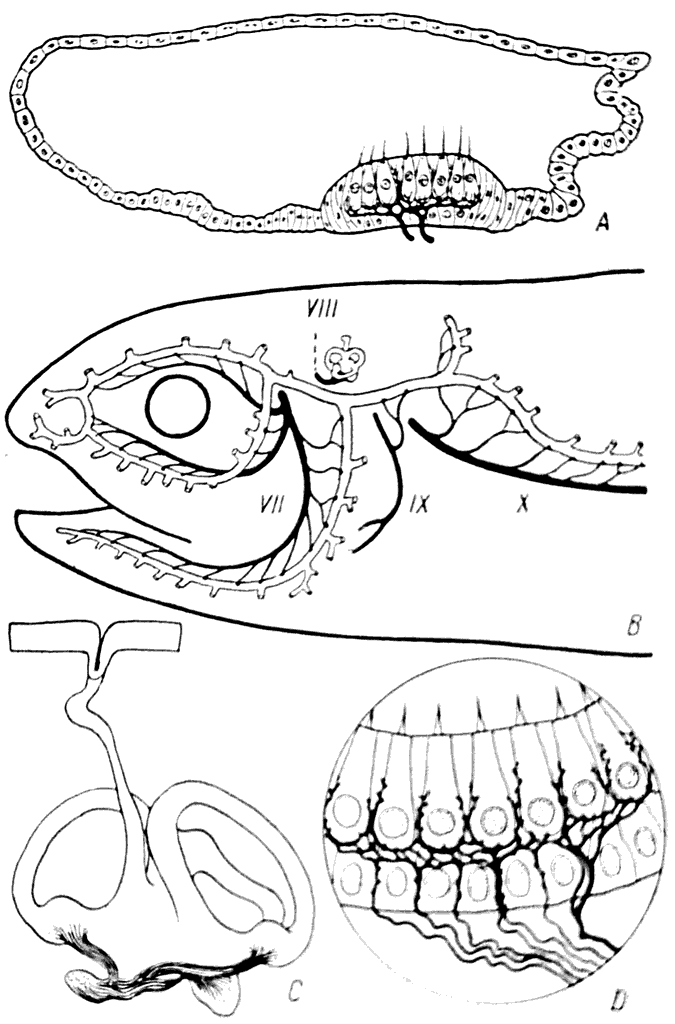
Рис. 31.
А — поперечный разрез сейсмосенсорного канала акулы (Mustelus canis); В — сейсмосенсорная система и ее иннервация у костистых рыб; С — перепончатый лабиринт акулы Scyllorhynchus; D — чувствительные клетки в перепончатом лабиринте мыши.
На голове рецепторы сейсмосенсорной системы иннервирутся ветвями nегvus facialis (VII) и на очень небольшом протяжении — n. glossopharyngeus (IX); на туловище — ветвями nervus vagus (X) (рис. 31, В).
Функция сейсмосенсорной системы до сих пор является для нас физиологической загадкой. Мы не можем ясно представить себе ее действие, так как у человека от нее не сохранилось никаких следов. Однако нужно думать, что рецепторные органы этой своеобразной системы реагируют на вибрационные раздражения.
Так как вся сейсмосенсорная система при помощи многочисленных отверстий сообщалась с внешней средой (с водой), то легко себе представить, что всякие колебательные движения в ней в той или иной мере приводили в движение волоски клеток сейсмосенсорных рецепторов и возбуждали их, что тотчас же по соответствующему нерву передавалось в головной мозг животиого. Если сейсмосенсорная система функционировала именно так, то она должна быть признана очень важной для всех водных форм позвоночных. Она позволяла им воспринимать разнообразные колебательные движения, например волны, вызванные движением других животных, и, может быть, даже слабые отраженные волны, возникающие в результате собственных перемещений позвоночных. Такая функция сейсмосенсорной системы позволяла водным позвоночным даже при слабом освещении легко ориентироваться в окружающей обстановке.
Кроме Agnatha, покрытых изолированными кожными зубами (Coelolepida) или снабженных панцирем, состоящим из сильно видоизмененных и сросшихся друг с другом зубов (Heterostraci), в силуре появились и такие формы бесчелюстных позвоночных, у которых все тело было покрыто несросшимися элементами наружного скелета, нигде не образующими сплошного щита.
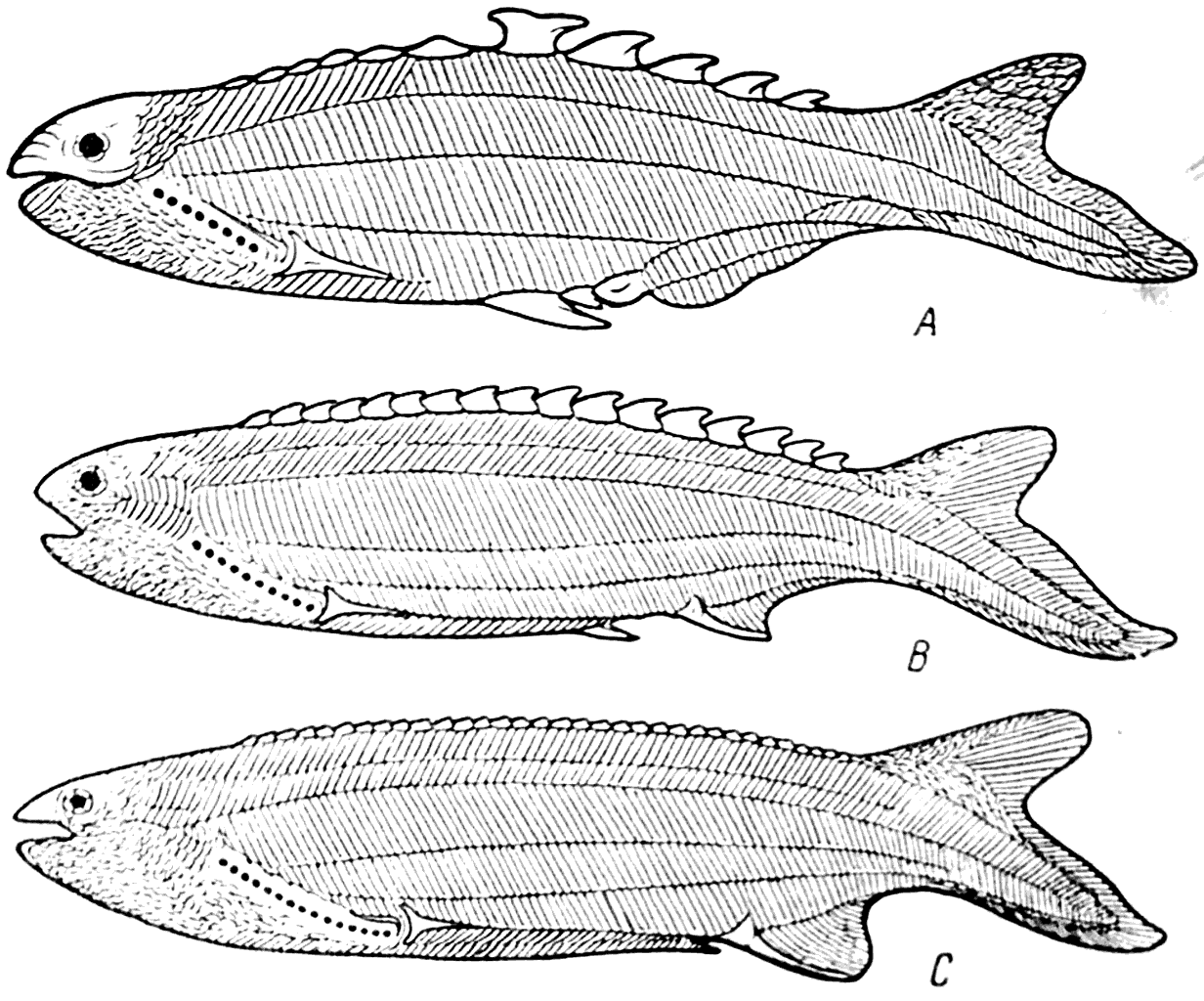
Рис. 32. Anaspida: Birkenia elegans (A), Pterolepis nitida (В), Pharyngolepis oblonga (С) (силур).
Эти представители древних бесчелюстных позвоночных, относящиеся к отряду Anaspida (рис. 17 — А), были небольшими рыбообразными формами, их длина колебалась от 2 до 20 см (рис. 32, A, B, C). Голова их была покрыта маленькими чешуйками, а туловище — несколькими рядами длинных узких пластинок. Глаза у Anapsida была хорошо развиты и располагались на боковых сторонах головы. Края беззубого рта были покрыты панцырными пластинками. На боковой стороне переднего отдела туловища располагались в виде косого ряда круглые жаберные отверстия; их число колебалось от 6 до 15. На спине у Anaspida вместо дорзальных плавников помещался ряд своеобразных шипов. Грудные и брюшные плавники у них отсутствовали; анальный плавник был более или менее развит. Хвост у всех Anaspida был гипоцеркальным.
Как показало изучение под микроскопом, маленькие панцырные элементы Anaspida не содержат ни дентинных канальцев, ни полостей костных клеток и, следовательно, состоят из аспидина; в нем удается обнаружить большое количество шарпеевских каналов (рис. 33).
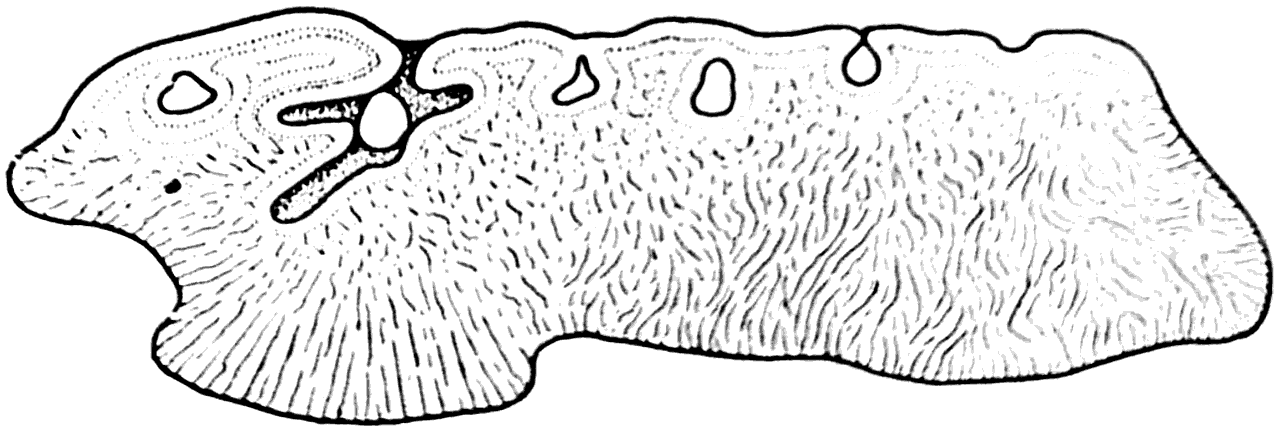
Рис. 33. Поперечный разрез чешуи одного из представителей анаспид (×300) (А. П. Быстров, 1955).
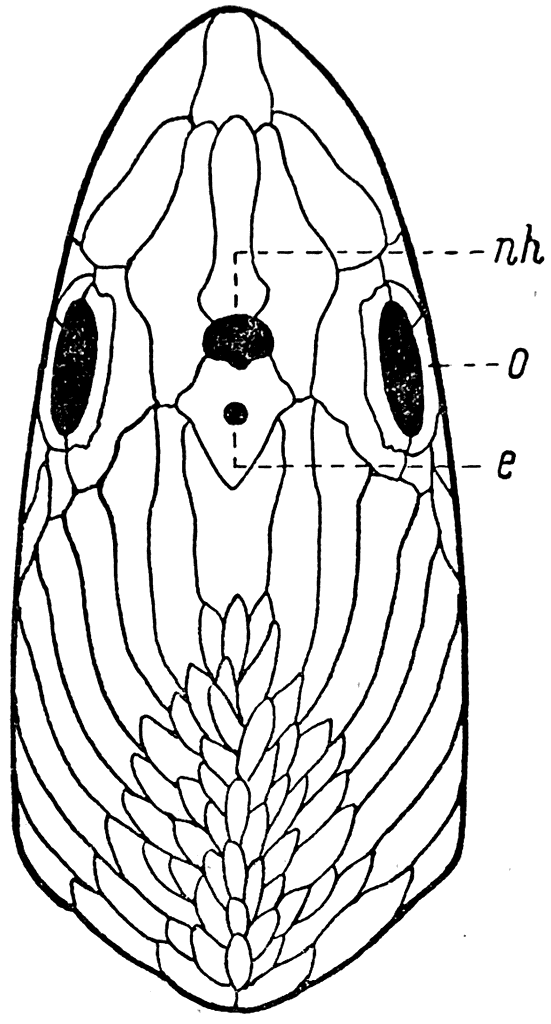
Рис. 34. Голова Pharyngolepis
oblonga (вид сверху).
е — отверстие для эпифиза; nh — отверстие для назогипофизарного органа; о — орбита.
Anaspida, по всей вероятности, произошли от каких-то примитивных и древних представителей целолепид. Эти целолепиды не были придонными формами; их тело было покрыто многочисленными кожными зубами, снабженными аспидиновыми отростками. Срастание этих отростков у первых анаспсид привело к образованию на их теле многочисленных коротких и длинных панцырных пластинок, состоящих из аспидина. Кожные зубы на поверхности этих панцырных элементов затем подверглись полной редукции. Все Anaspida, несомненно, были очень подвижными животными. Будучи способными быстро плавать, они, по всей вероятности, относительно легко избегали нападений преследующих их врагов. Этим, видимо, и объясняется отсутствие у них такого хорошо развитого панцыря, какой имелся у менее подвижных Heterostraci.
На верхней поверхности головы у Anaspida между их большими орбитами располагались два непарных отверстия (рис. 34). В заднем меньшем по своим размерам отверстии помещался пинеальный орган (эпифиз мозга — epiphysis cerebri). Что же касается большего переднего отверстия, то назначение его, вероятно, навсегда осталось бы для нас непонятным, если бы мы не имели возможности изучить строение и развитие так называемого назогипофизарного органа у современных миног.
У личинки миноги между ротовой бухтой (рис. 35, А — m) и обонятельными ямками образуется так называемый гипофизарный мешок (h). Его вершина направлена косо вверх и назад к переднему концу хорды (ch). По мере развития миноги происходит сильное разрастание того участка эктодермы, который располагается между гипофизарным мешком и ротовой бухтой. Это в конце концов приводит к тому, что гипофизарный мешок (h) и обонятельные ямки (n) смещаются с вентральной стороны головы на дорзальную и у взрослого животного оказываются расположенными следующим образом. Конец гипофизарного мешка (h) лежит непосредственно под передним концом хорды (ch); его выводной проток идет косо вверх и вперед, а сближенные обонятельные ямки (n) открываются в него сзади. Так формируется и так построен назогипофизарный орган у современных миног.
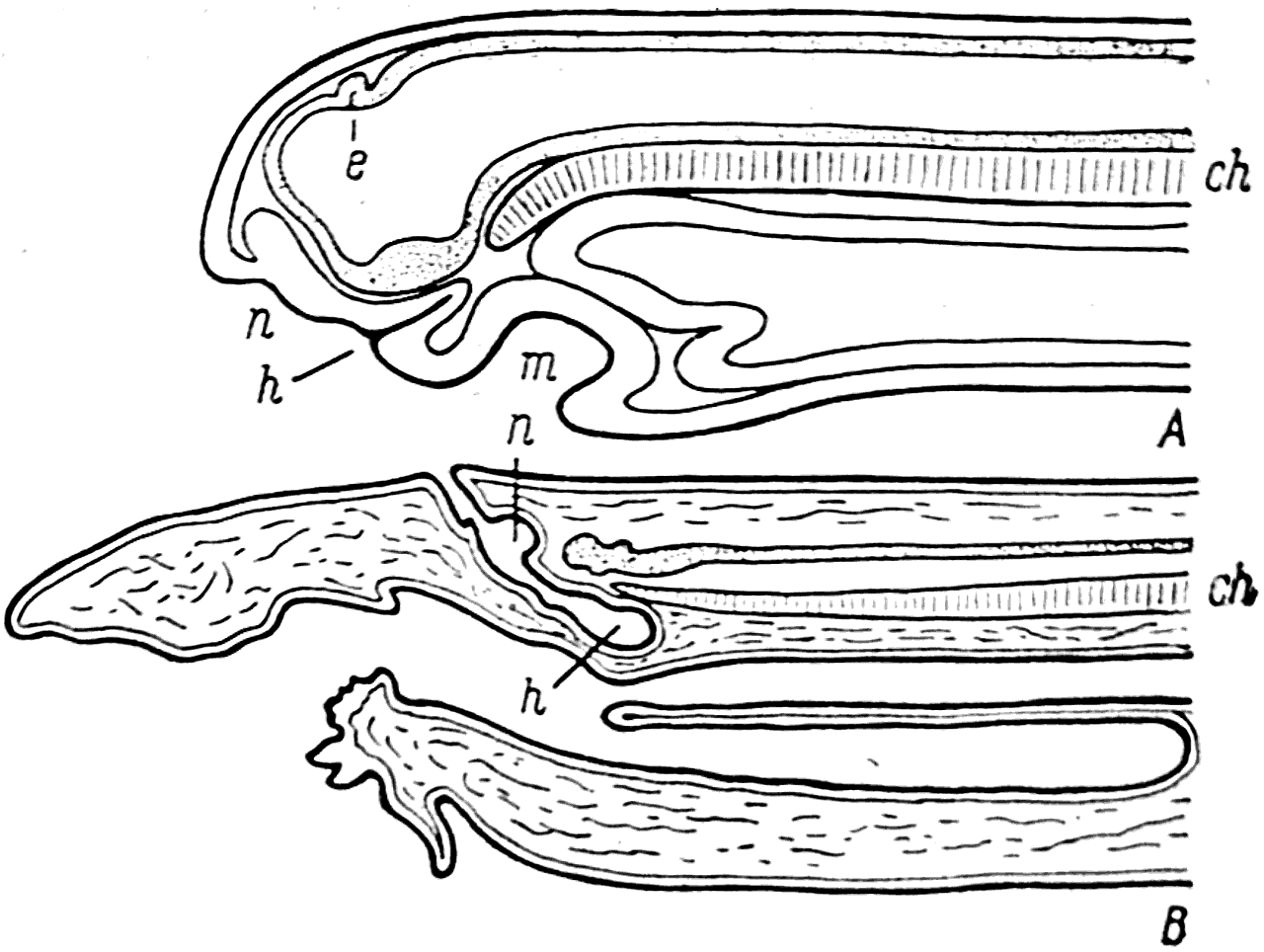
Рис. 35.
А — продольный разрез головы личинки миноги; В — такой же разрез головы взрослой миноги.
ch — хорда; е — эпифиз; h — гипофиз; n — обонятельная ямка; m — ротовая бухта.
У Anaspida строение назогипофизарного органа, несомненно, было таким же, и он открывался у них на верхней стороне головы через отверстие, расположенное кпереди от foramen pineale (рис. 34 — nh).
В отличие от Heterostraci и Anaspida силурийские Agnatha, относящиеся к отряду Osteostraci, были снабжены панцырем, состоящим из настоящей кости. Эти бесчелюстные позвоночные по всей вероятности являлись потомками анаспид (рис. 17 — O). Представители Osteostraci были сходны с Anaspida тем, что также имели хорошо развитый назогипофизарный орган. Но все они отличались от Anaspida тем, что были типичными придонными формами и по своему виду напоминали скатов.
Костный панцырь Osteostraci, по-видимому, образовался из аспидиновых панцырных элементов анаспид вследствие того, что соединительно-тканные клетки стали замуровываться в образованный ими аспидин и тем самым превратили его в кость.
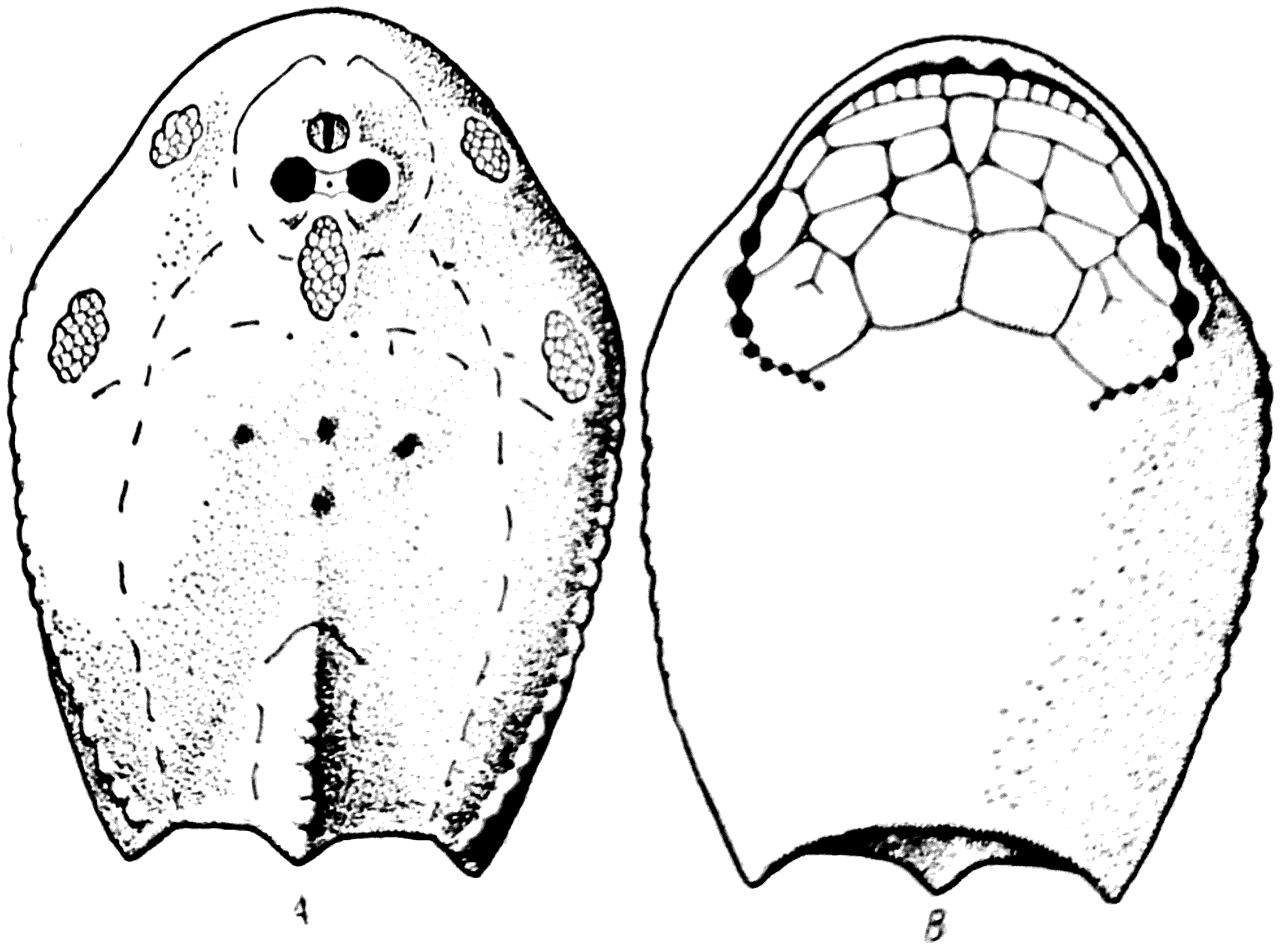
Рис. 36. Tremataspis schmidti (силур). А — вид сверху; В — вид снизу.
Одна из форм Osteostraci — Tremataspis schmidti (рис. 36), имела сильно уплощенную голову, сверху и снизу покрытую панцырными щитами, сросшимися по краям. Длина панцыря равнялась 3,5 см, ширина — 2,5 см. На верхней стороне головного щита Tremataspis можно видеть сближенные друг с другом орбиты, между которыми располагается небольшое пинеальное отверстие (рис. 36, А). Кпереди от этого отверстия лежит еще одно непарное отверстие, служившее местом выхода назогипофизарного органа.
Позади орбит в сплошном панцыре Tremataspis имеется непарный дефект, заполненный мелкими изолированными костными элементами. Две пары таких же дефектов помещаются справа и слева неподалеку от боковых краев головного щита.
Детальное изучение некоторых Osteostraci дало возможность Стеншио (Stensiö, 1927) высказать предположение, что под этими полями, состоящими из мелких костных элементов, у остеостраков располагались электрические органы.
Позади центрального электрического поля у Tremataspis имеются два маленьких отверстия, которые служили местом выхода ductus endolymphaticus на поверхность кожи животного.
Изучение развития перепончатого лабиринта у современных позвоночных показало, что он образуется в результате впячивания эктодермы в виде кармана в область слуховой капсулы хрящевого черепа (рис. 37, A, В, С, D).
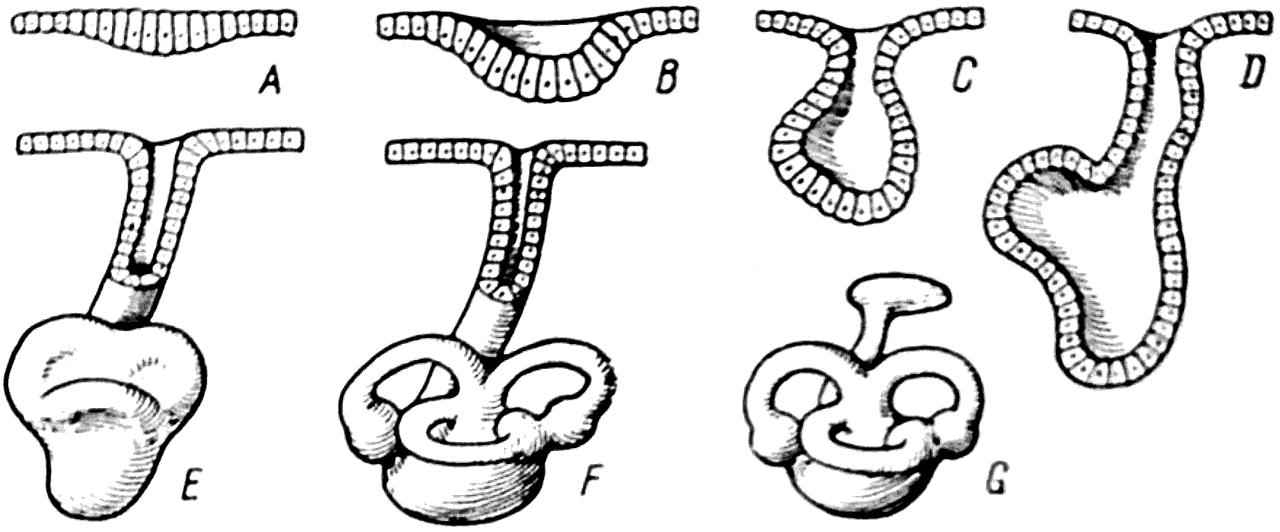
Рис. 37. Последовательные фазы развития перепончатого лабиринта (схема).
Из этого эктодермального мешка затем формируются перепончатые полукружные каналы и другие отделы внутреннего уха. На начальных стадиях своего развития лабиринт еще сохраняет сообщение с наружной средой при помощи так называемого ductus endolymphaticus (рис. 37, Е, F), но впоследствии этот проток у большинства позвоночных закрывается и лабиринт внутреннего уха совершенно изолируется (рис. 37, G).
Присутствие пары маленьких отверстий на щите Tremataspis свидетельствует о том, что полости его перепончатых лабиринтов еще сообщались с внешней средой. Интересно, что Стеншио (1927) удалось показать, что лабиринт у представителя Osteostraci — Kioeraspis, имел только два полукружных канала — передний и задний (рис. 38, А); латеральный канал отсутствовал. Такую же особенность мы наблюдаем и у миноги (рис. 38, B).
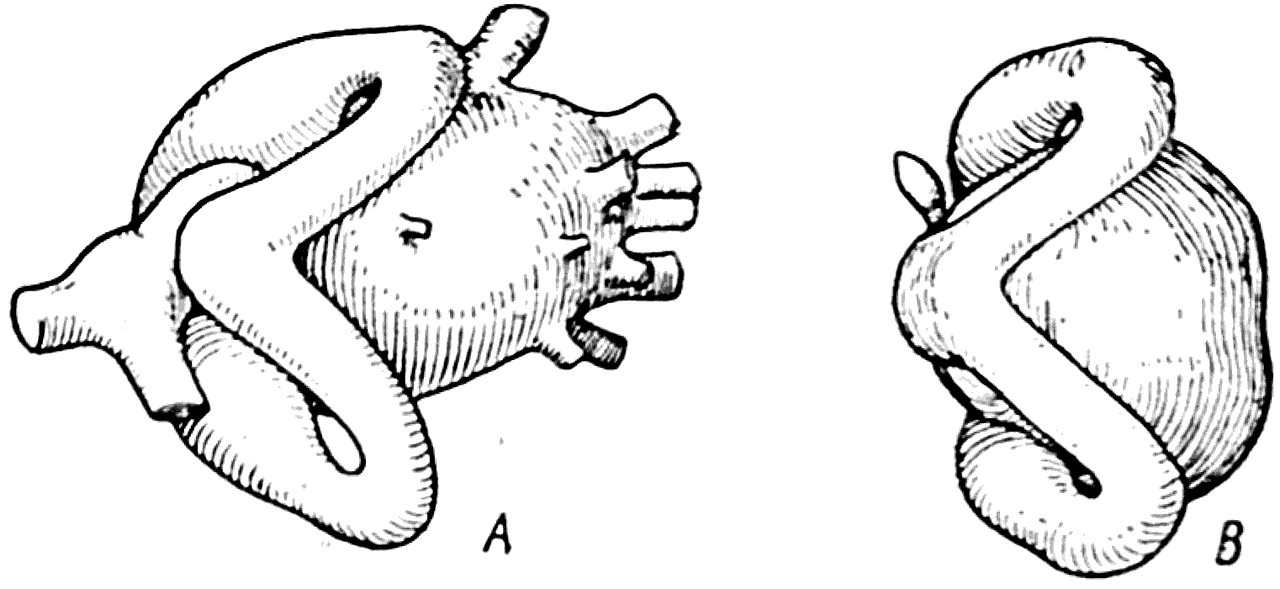
Рис. 38.
А — левый лабиринт одного из представителей Osteostraci (Kiaeraspis, Стеншио, 1927); В — левый лабиринт миноги (вид сверху).
В нашем распоряжении нет прямых морфологических доказательств того, что все без исключения силурийские (и девонские) Agnatha имели в своем перепончатом лабиринте два полукружных канала, но у нас нет оснований и отрицать это.
Интересно, что на перепончатый лабиринт позвоночных мы можем смотреть как на образование, по своему происхождению связанное с их сейсмосенсорной системой. Он возник, во всей вероятности, в результате погружения части сейсмосенсорной системы в полость слуховой капсулы хрящевого черепа. Доказательством этого являются следующие факты.
Лабиринт развивается, как и сейсмосенсорная система, из эктодермы, и на первых фазах формирования его полость сообщается с внешней средой. Таким образом, ductus endolymphaticus можно считать гомологичным тем трубочкам, при помощи которых сейсмосенсорные каналы открываются на поверхности кожи. (У некоторых акул это сообщение лабиринта сохраняется в течение всей жизни, рис. 31, C)
Некоторые участки эпителиальной стенки лабиринта имеют такие же клетки с волосками, какие находятся и в рецепторных аппаратах сейсмосенсорной системы (рис. 31, D).
Построенные таким образом чувствительные поля в перепончатом лабиринте у всех позвоночных иннервируются ветвями n. statoacusticus (VIII). Этот нерв выходит из мозга между n. facialis (VII) и n. glossopharyngeus (IX).
Все это дает нам право предполагать, что в процессе развития позвоночных некоторый участок сейсмосенсорного канала отделился от всей остальной системы, погрузился в хрящевой череп и здесь превратился в лабиринт. Это привело к тому, что нерв, выходящий из мозга между n. facialis (VII) и n. glossopharyngeus (IX) и иннервирующий этот участок, превратился в n. statoacusticus (VIII). Таким образом, сохраняя по-прежнему способность реагировать на колебательные раздражения, часть сейсмосенсорного канала после очень больших анатомических изменений превратилась в перепончатый лабиринт — орган слуха и сохранения равновесия.
На нижней стороне панцыря Tremataspis можно видеть два ряда жаберных отверстий (рис. 36, В). Передний отдел вентральной части щита у этого животного состоит из отдельных кусков панцыря. Отсутствие сращений между ними позволяло животному опускать и поднимать дно рта во время дыхательных движений и при захватывании пищи. Рот у Tremataspis представлял собой узкую поперечную щель, ограниченную спереди краем головного щита, а сзади — рядом мелких костных пластинок, которые приводились в движение при захватывании пищи.
Толщина панцыря Tremataspis была равна 0,3 мм. При изучении строения панцыря под микроскопом в его веществе легко удается обнаружить полости остеоцитов, соединяющиеся друг с другом тонкими канальцами (рис. 39). Следовательно, панцырь Tremataspis состоял из костной ткани.
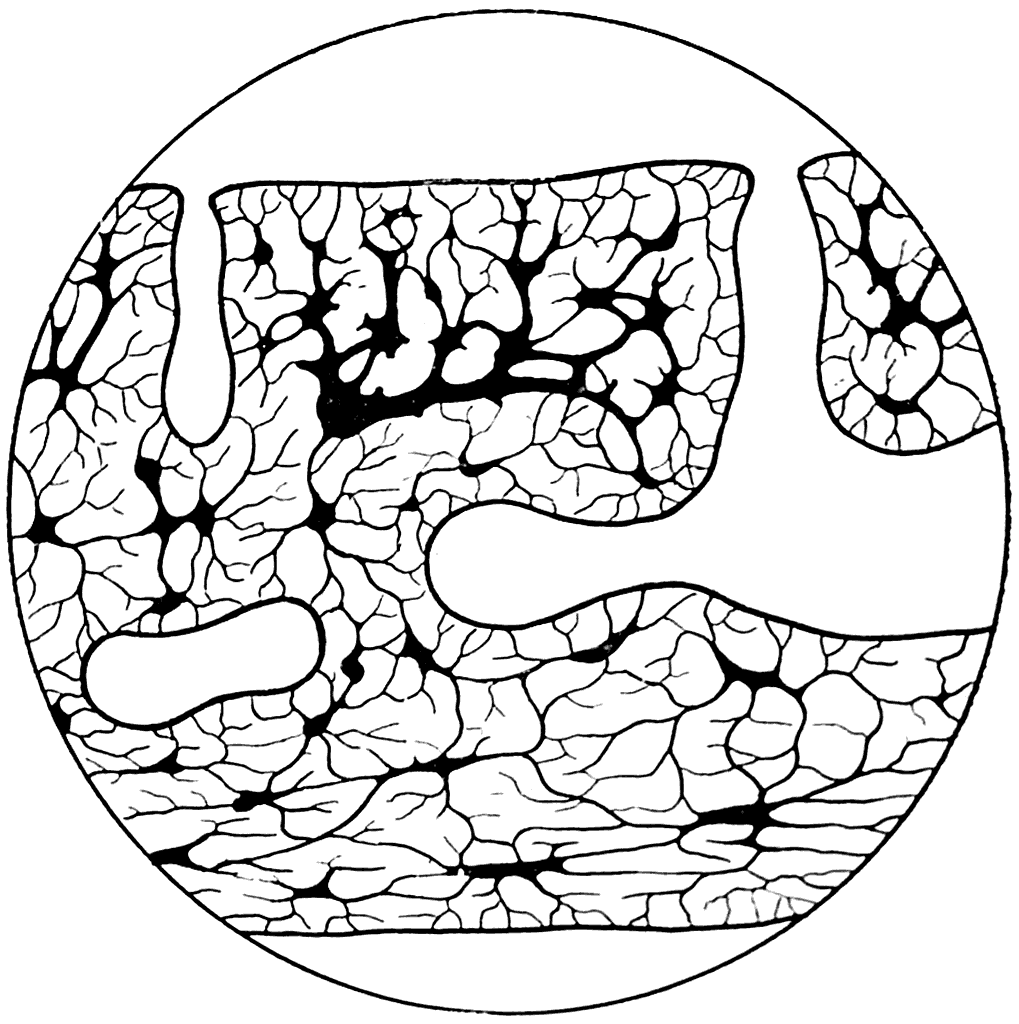
Рис. 39. Вертикальный разрез панцыря Tremataspis schmidti (×300) (А. П. Быстров, 1955).
В среднем слое панцыря имеются довольно обширные полости, от которых отходят вертикальные каналы, открывающиеся на поверхности щита отверстиями или порами. Эти поры хорошо видны при рассматривании поверхности панцыря под лупой.
Костный панцырь силурийских Osteostraci, несомненно, не был вполне надежной защитой от врагов, так как его толщина колебалась от 0,1 до 0,3 мм. Нужно думать, что именно это обстоятельство было причиной того, что у всех представителей этой группы Agnatha возникли электрические органы, способные своим разрядом оглушить нападающего врага. Таким образом, наличие электрических органов следует рассматривать как компенсацию недостаточной прочности панцыря.
Тот факт, что сближенные глаза у Osteostraci располагались на дорзальной стороне головного щита, является несомненным доказательством того, что эти своеобразные представители Agnatha, прячась от своих врагов, обычно зарывались в ил на дне водного бассейна и там занимались поиском своей пищи. При этом те хищники, которые случайно наталкивались на них, получали такой электрический удар, который, вероятно, был способен заставить их прекратить преследование.
Детальное изучение всех известных нам силурийских Agnatha показывает, что ни один из представителей этого класса не может рассматриваться в качестве возможного предка позвоночных, снабженных челюстным аппаратом (Gnathostomata). Все эти формы были в той или иной степени специализированными животными.
Большинство силурийских целолепид было придонными формами, и их жизнь на дне сильно отразилась на их строении.
Heterostraci, снабженные хорошо развитым панцырем, состоящим из сросшихся и сильно видоизмененных кожных зубов, уже не могли дать лишенного панцыря предка Gnathostomata.
Наличие назогипофизарного органа у Anaspida не дает нам права искать среди них предков рыб, у которых, как известно, перемещения гипофизарного кармана (карман Ратке) на дорзальную сторону головы никогда не происходит.
Появление у Osteostraci костной ткани нельзя рассматривать как указание на их близость к рыбам. Эти Agnatha, как и Anaspida, имели назогипофизарный орган и, следовательно, предками примитивных рыб быть не могли.
Кроме того, следует иметь в виду, что остатки примитивных Gnathostomata найдены в силурийских отложениях. Следовательно, они. были современниками многих древних форм Agnatha.
Все это свидетельствует о том, что непосредственных предков Gnathostomata следует искать где-то среди ордовикских неспециализированных бесчелюстных позвоночных. Но, к сожалению, о них мы знаем очень мало и, возможно, никогда не будем знать достаточно. Если в скелете этих форм твердыми образованиями, как у Palaeodus, были только кожные зубы, игравшие роль чешуи, то в таком случае мы не имеем оснований надеяться когда-нибудь найти в нижнесилурийских отложениях что-нибудь более ценное, чем рассыпанные в породе кожные зубы.
Быстров А. П. Микроструктура панцыря бесчелюстных позвоночных силура и девона. Сборник памяти акад. Л. С. Берга, изд. АН СССР, 1955.
Кlanег I. and Неintz A. The Downtonian and Devonian vertebrates of Spitsbergen. Skrifter om Svalbard og Ishavet, № 40, 1935.
Pander Ch. Monographie der fossilen Fische des Silurischen Systems. St-Petersburg, 1856.
Rоhоn I. W. Über untersilurischen Fische. Melanges Gèologiques et paleontologiqucs Theses du Bulletin de l’Academie Imperial des Sciences. Т. I, St. Petersbourg, 1889.
Stensiö E. The Downtonian and Devonian vertebrates of Spitsbergen. Scrifter om Svalbard og Nordishavet, N 12, 1927.