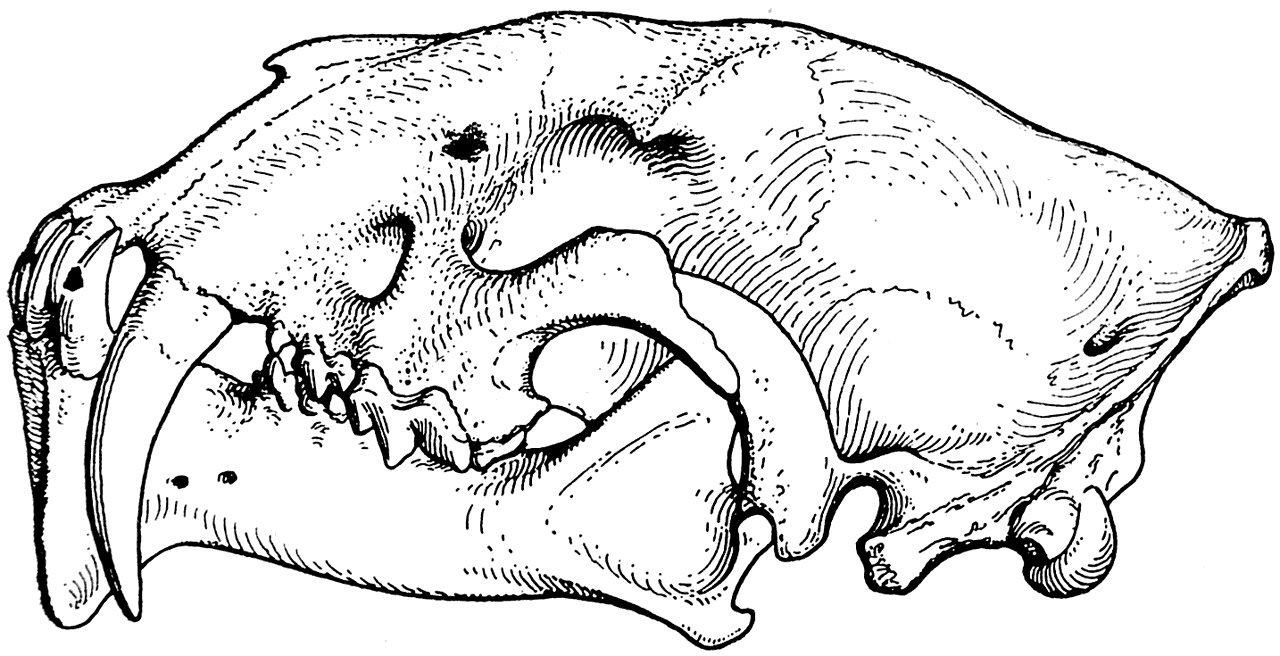
Рис. 6. Череп Drepanodon primaevus. Северная Америка, олигоцен (Скотт и Джепсен, 1936).
Как известно, Ч. Дарвин придавал большое значение данным палеонтологии, которые позволяют проследить изменения организации живых существ на протяжении многих миллионов лет. Но в 1859 г. он имел все основания жаловаться на «неполноту геологических летописей».
После выхода в свет книги Ч. Дарвина о происхождении видов в течение почти столетия в результате усиленных поисков палеонтологи накопили очень большой материал по истории позвоночных. Огромная теоретическая ценность этого материала, разумеется, не подлежит никакому сомнению. Однако, непрерывно пополняя соответствующие музеи все новыми и новыми находками, палеонтологи получают возможность изучать только фактическую сторону эволюционного процесса. В некоторых случаях ряд ценных находок позволяет им выяснить основные этапы филогенетического развития той или иной группы животных. Но при этом исследователи обычно не находят никаких указаний на те конкретные причины, которые заставили данное животное пройти именно такой, а не иной путь эволюционного развития.
В отличие от палеонтологов зоологи, изучающие современных животных, имеют возможность выяснить не только все их особенности как живых организмов, но и исследовать те сложные взаимоотношения, которые обусловливают их эволюционное развитие. Однако зоологи не в состоянии непосредственно наблюдать ход эволюционного процесса.
Таким образом, изучение прошлого привело к констатации эволюции без выяснения ее факторов, а изучение настоящего привело к выяснению факторов эволюции без достаточно очевидной ее констатации.
Такое положение потребовало допущения, что при изменениях древних животных и растений действовали те же факторы эволюции, которые действуют и в современном органическом мире, и что эволюционный процесс, констатированный для прошлого, продолжается и в настоящее время. Иными словами, в прошлом мы констатируем факты эволюции, а в настоящем мы констатируем факторы эволюции; в прошлом мы предполагаем факторы эволюции, в настоящем мы предполагаем факты эволюции.
Все это приводит к тому, что одни палеонтологи верят, другие допускают, третьи отрицают, что причинами эволюционных изменений являются факторы, установленные Ч. Дарвином. Так как эти факторы пока еще не могут быть доказаны ископаемым материалом, то именно среди палеонтологов чаще всего встречаются противники теории Дарвина. Не имея возможности проследить влияние дарвиновских факторов на филогенетические изменения вымерших форм, антидарвинисты обычно утверждают, что теория Дарвина неверна. Для ее замены они нередко создают, хотя и совершенно неубедительные, но новые гипотезы.
Впрочем, не все антидарвинисты полностью отвергают теорию Дарвина. Некоторые из них пытаются внести в нее ряд дополнений и исправлений. Без таких дополнений дарвинизм, по их мнению, не может объяснить многие факты из истории развития живых существ. Но все эти дополнения и исправления не улучшают и не уточняют теорию Дарвина, а превращают ее в совершенно ненаучную гипотезу. Наиболее ярким примером теории такого типа следует признать теорию Додерлейна (L. Döderlein, 1888).
В качестве основного примера, которым Додерлейн иллюстрирует свою теорию, является история эволюции вымерших саблезубых тигров.
Судя по довольно богатому ископаемому материалу, эти тигры прошли очень своеобразный путь эволюционного развития, на протяжении которого у них постепенно увеличивались размеры верхних клыков и шла редукция коренных зубов. Одновременно с этим произошли некоторые изменения и в челюстных суставах (рис. 6 и 7; Скотт и Джепсен — Scott and Jepsen, 1936).
Рис. 6. Череп Drepanodon primaevus. Северная Америка, олигоцен (Скотт и Джепсен, 1936).
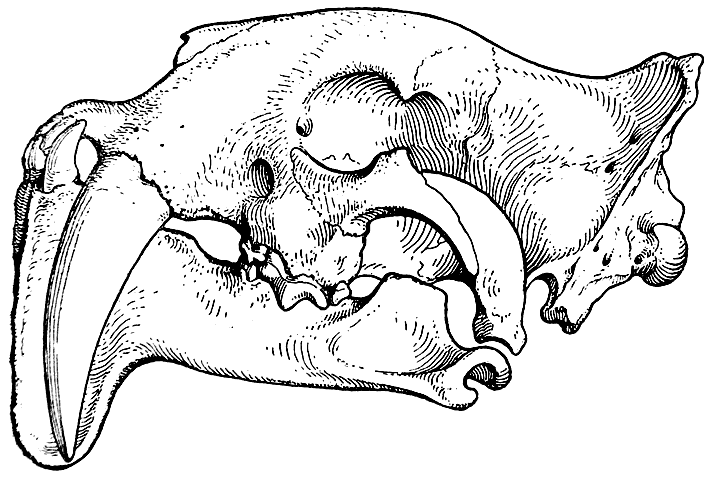
Рис. 7. Череп Eusmilus sicarius. Северная Америка, олигоцен (Скотт и Джепсен, 1936).
Л. Додерлейн согласен с тем, что образование больших клыков на верхних челюстях у саблезубых тигров было вызвано естественным отбором, закрепляющим полезные варианты строения. Но при этом постепенное увеличение клыков под влиянием отбора, по его мнению, продолжалось до тех пор, пока эти мощные зубы не дали тиграм возможности вытеснить и полностью истребить всех своих конкурентов.
«Можно и даже должно считать, — говорит Додерлейн, — что если у ряда животных важные органы, служащие, например, оружием, достигают высшей целесообразности, то это настолько вредит их прежним конкурентам, что они уступают им место».
Таким образом, сильное развитие клыков у саблезубых тигров, по мнению Додерлейна, привело к тому, что борьба за существование сначала значительно ослабла для них, а затем совсем исчезла. Одновременно с этим, разумеется, должен был прекратить свое действие и естественный отбор. Это, казалось бы, должно было повлечь за собой и немедленную остановку дальнейшей эволюции зубов. Однако Додерлейн полагает, что дальнейшее развитие клыков и после этого пррдолжалось, но теперь уже только под влиянием одной «силы инерции».
«Здесь имеется своего рода vis inertiae» — говорит он. Таким образом, по Додерлейну, естественный отбор обусловливал эволюционный процесс только до момента возникновения у животного максимально целесообразного строения органа. Все же последующее развитие совершалось только по инерции, причем, направление развития в этом случае диктовалось уже пройденным эволюционным путем.
«Я считаю возможным думать, — пишет Додерлейн, — что после того, как у длинного ряда предков упорно сохранялось определенное направление развития, пока, например, не достигалось состояние наибольшей целесообразности, потомки наследуют не это определенное состояние, а только тенденцию развиваться именно в этом определенном направлении».
Следовательно, по мнению Додерлейна, развитие под влиянием «силы инерции» являлось не только непосредственным продолжением развития, протекавшего под действием факторов Ч. Дарвина, но и совершалось оно в том же направлении, в котором шло прежде. В результате этого клыки животного продолжали увеличиваться. Это увеличение с некоторого момента было уже ненужным и нецелесообразным. В первое время при отсутствии уже уничтоженных конкурентов, а следовательно, и при отсутствии борьбы за существование и естественного отбора, большие, но бесполезные клыки не мешали хищнику жить. Но потом дальнейшее увеличение этих зубов привело к такой их длине, что животное лишилось возможности есть (рис. 8).
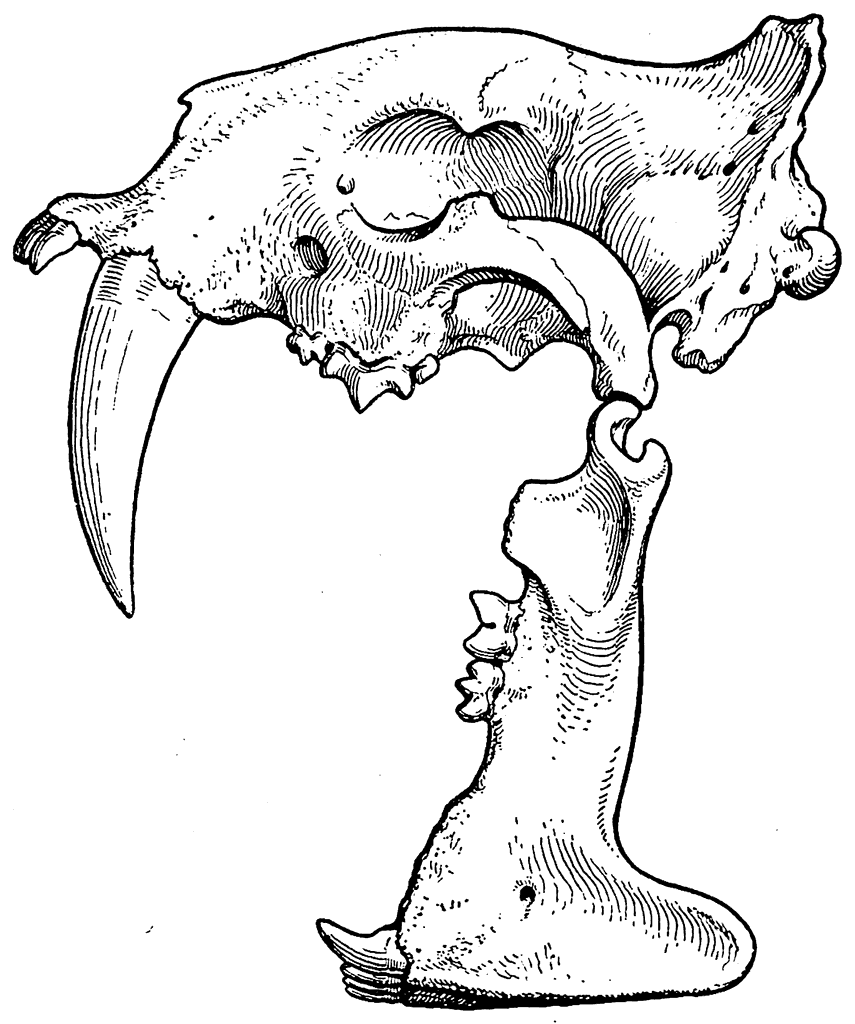
Рис. 8. Череп Eusmilus sicarius (рот открыт).
«Эти зубы, — говорит Додерлейн, — сами по себе представлявшие ужасное оружие, одновременно с этим вследствие своей необыкновенной длины чрезвычайно мешали при еде, действуя как намордник». Это обстоятельство, по мнению Додерлейна, и было причиной вымирания саблезубых тигров.
Таким образом, эволюционная сила инерции или унаследованная тенденция развития в конце концов привела этих своеобразных хищников к гибели.
Такие мощные клыки, как у наиболее специализированных саблезубых тигров (Eusmilus, рис. 7), были обнаружены и у найденного в 1926 г. в плиоценовых отложениях Аргентины сумчатого хищника Thylacosmilus atrox (рис. 9; Риггс — Riggs, 1933). Если бы это своеобразное животное было найдено лет на 40 раньше, то Л. Додерлейн, несомненно, воспользовался бы им в качестве хорошей иллюстрации своей гипотезы.
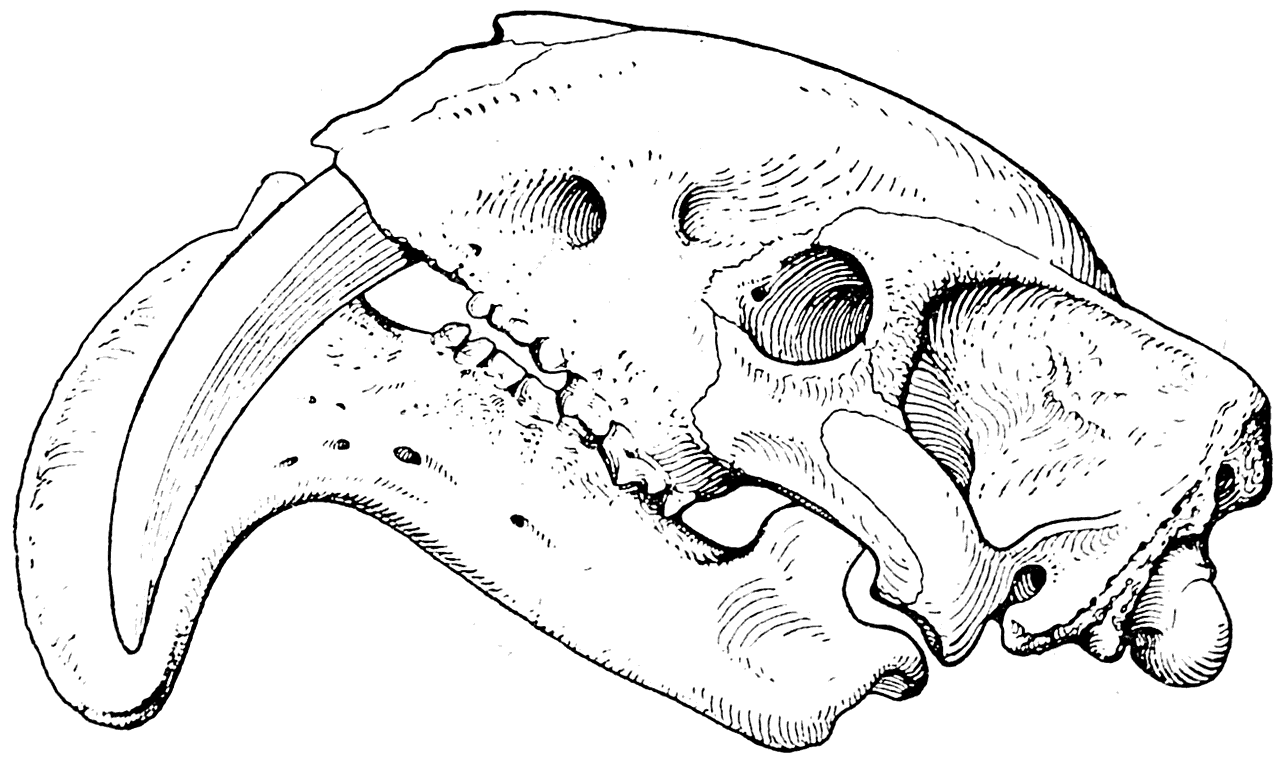
Рис. 9. Череп Thylacosmilus atrox. Сумчатый хищник из плиоцена Аргентины (Риггс, 1933).
Мы не знаем, какими животными питался Thylacosmilus atrox и какую пользу приносили ему его огромные клыки. Однако можно уверенно сказать, что они для него, несомненно, представляли большую ценность. Доказательством этого является то, что подбородочная часть нижней челюсти животного сильно разрослась вниз и, помещаясь при закрытом рте между клыками, предохраняла их от случайных переломов. Такое же приспособление, как известно, имелось и у многих саблезубых тигров, например у Drepanodon (рис. 6), и было сильно развито у Eusmilus (рис. 7).
Кроме ссылки на эволюцию саблезубых тигров, Л. Додерлейн в подтверждение своей гипотезы указывает на огромные бивни мамонта (рис. 10) и на необычайно сильное развитие рогов у плейстоценового оленя Megaceros (рис. 11).
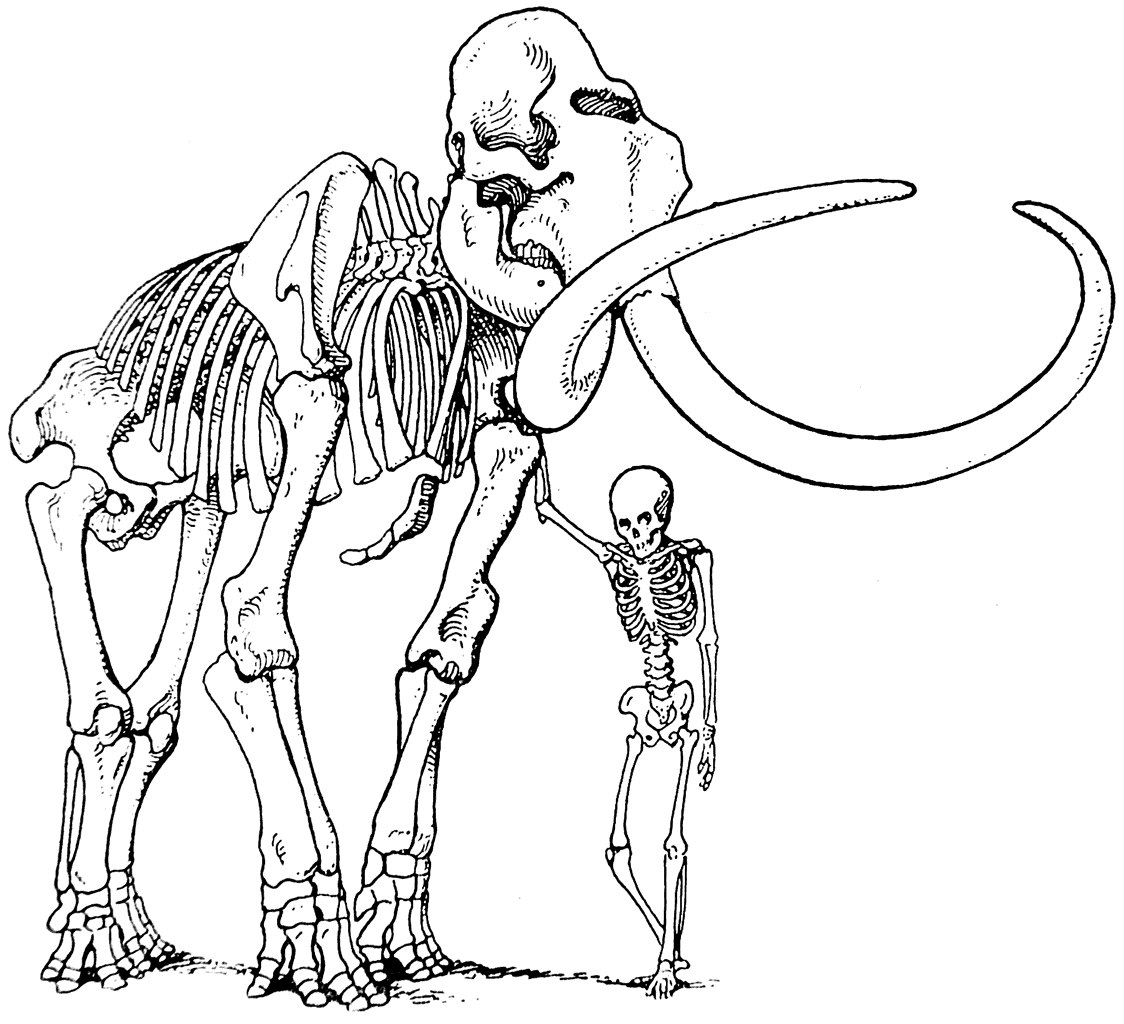
Рис. 10. Скелет мамонта.
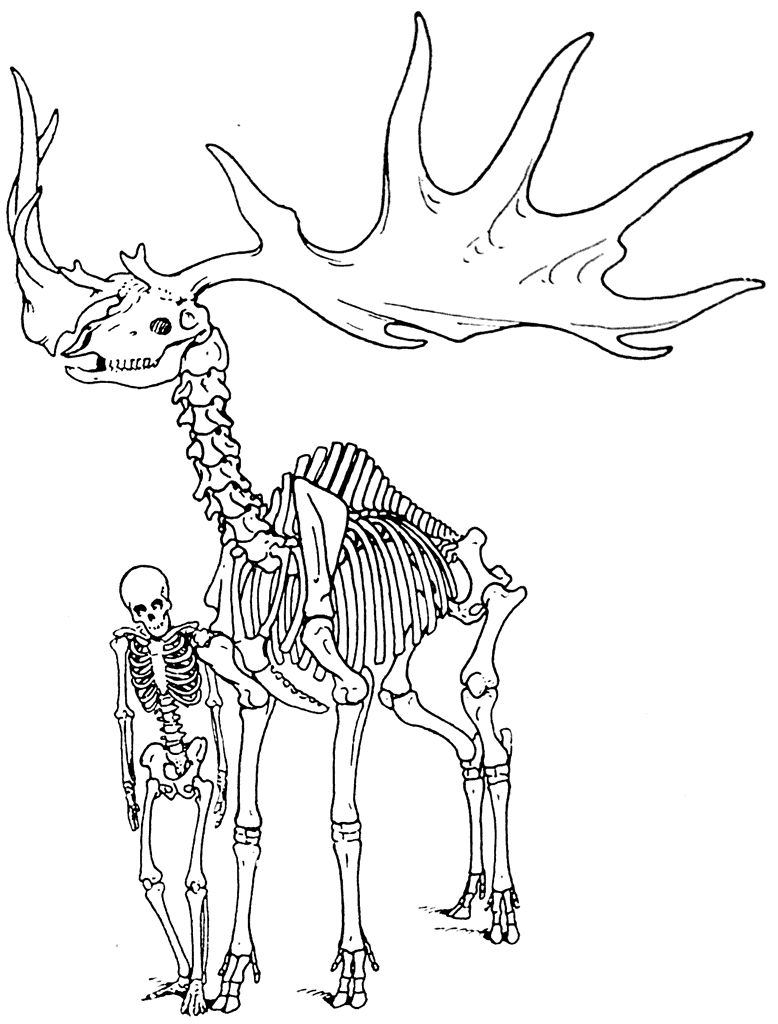
Рис. 11. Скелет Megaceros hibernicus Owen из торфяных болот Ирландии.
По мнению Л. Додерлейна, бивни мамонта и рога оленя достигали таких больших размеров в силу тенденции к увеличению. Эта тенденция была выработана предками мамонта и предками оленя. Л. Додерлейн считает, что в течение длительного периода эволюционного развития увеличение размеров бивней было полезно предкам мамонта, полезно было и увеличение рогов предкам оленя. Но после достижения состояния наибольшей целесообразности дальнейший рост и бивней и рогов был уже не только бесполезным, но даже вредным. Однако он продолжался до тех пор, пока не оказался причиной вымирания и мамонта и гигантского оленя.
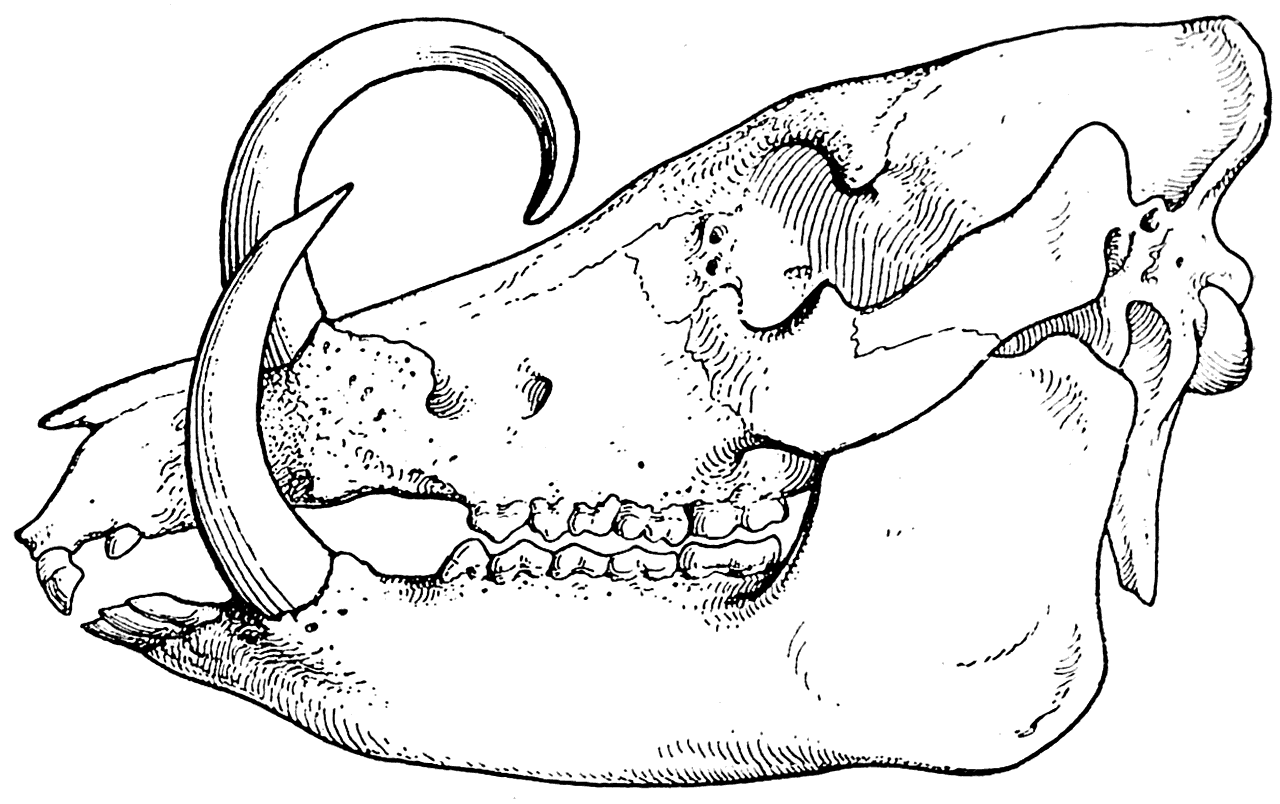
Рис. 12. Череп бабируссы (Babirussa babirussa).
Наконец, для доказательства правоты своих взглядов Л. Додерлейн указывает на сильное развитие верхних и нижних клыков у современной дикой свиньи бабируссы (Babirussa babirussa), живущей на острове Целебес (рис. 12). Он считает, что после того, как бабирусса была изолирована от крупных хищников на острове и перестала подвергаться нападениям, ее клыки продолжали развиваться в прежнем направлении только под влиянием vis inertiae и скоро достигли настолько нецелесообразных размеров, что в некоторых случаях стали своими концами врастать в череп (рис. 13).
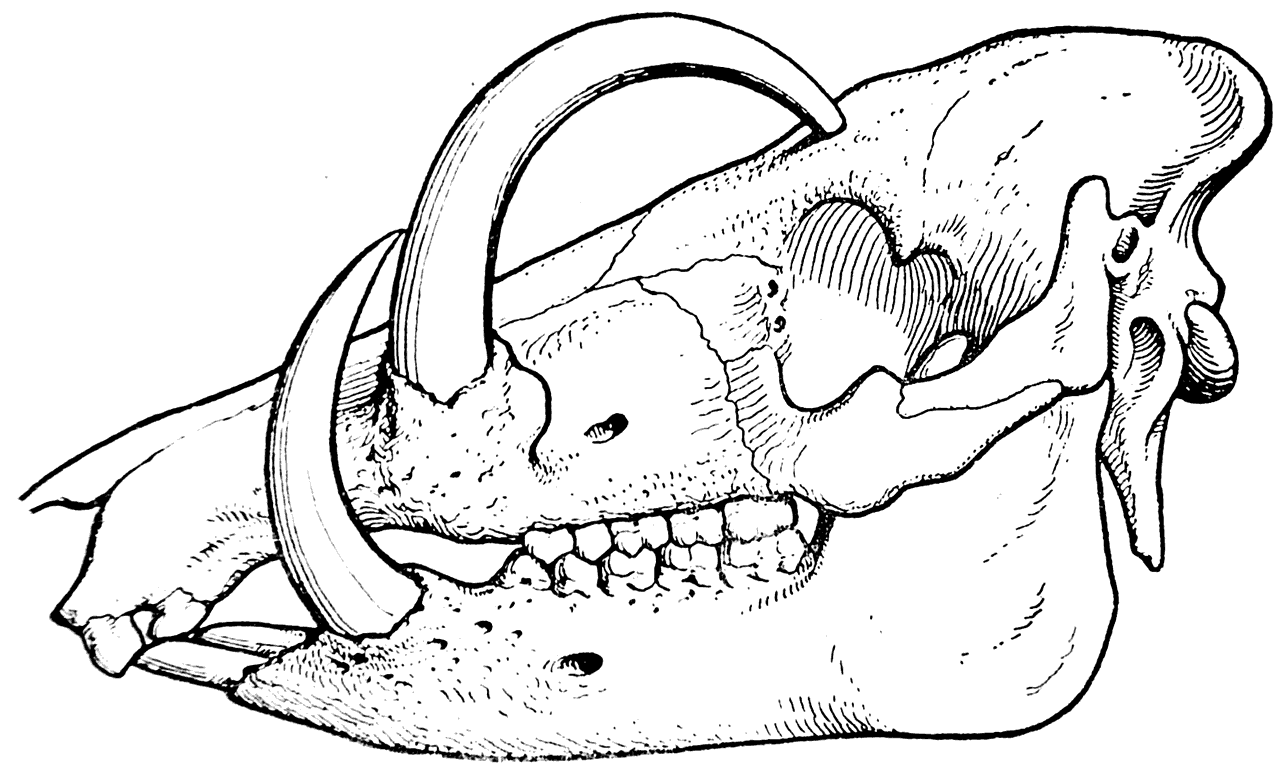
Рис. 13. Череп бабируссы (Babirussa babirussa).
Изучение гипотезы Додерлейна показывает, что его vis inertiae представляет собой некое дополнение к теории Дарвина, такое дополнение, без которого будто бы совершенно невозможно объяснить некоторые факты из истории эволюционного развития животных. По существу, все новое в гипотезе Л. Додерлейна сводится к утверждению, что эволюционные изменения являются в значительной степени внутренним свойством живых организмов, с чем нельзя согласиться.
В 1897 г. Т. Эймер (Th. Eimer) выдвинул типично антидарвиновскую теорию прямого развития, или «ортогенеза» (ὀρθός — orthos — прямой; γένεσις — genesis — происхождение). «Именно в силу того, — говорит он, — что органическое образование формы основывается на физико-химических процессах, оно является, как и форма неорганических кристаллов, определенным и при всяком новообразовании может идти только по определенным направлениям».
Из этого можно сделать вывод, что orthogenesis как эволюционный процесс является одним из свойств живых организмов. Однако, по мнению Эймера, «внешние воздействия, т. е. климат, свет, тепло, влажность, и различия в питании изменяют организмы без какого-либо участия отбора, и так как возникающие этим путем изменения наследующиеся, то они обусловливают возникновение новых видов».
«Отбор, — говорит Эймер, — не может создать решительно ничего нового».
Таким образом, эволюционный процесс, по Эймеру, оказывается обусловленным в значительной мере тем, что «плазма имеет свойство, будучи благодаря воздействию внешних сил измененной в физиологическом или в морфологическом отношении, принимать иной вид».
Следовательно, основной причиной появления изменений в плазме Т. Эймер считает внешние воздействия. Именно они (и только они) способны так или иначе изменять плазму живых организмов. Но эволюционные изменения вызываются не одним тем фактом, что плазма под влиянием внешних воздействий изменяется, а тем, что последовательные фазы этих изменений обусловливают соответствующую последовательность в этапах эволюционного развития организмов.
«Согласно моим исследованиям,— говорит Т. Эймер, — главной причиной эволюции является обусловленный внешними влияниями, климатом и пищей органический рост в плазме, выражением которого… является определенно направленное развитие (ортогенез)».
Таким образом, основным фактором эволюционных изменений Эймер считает прямое воздействие внешней среды, вызывающее в плазме такие изменения, которые приводят к направленной эволюции.
В 1922 г. Л. С. Берг опубликовал свою теорию «номогенеза» (νόμος — nomos — закон; γένεσις — genesis — происхождение). Он утверждал, что «эволюция организмов есть результат некоторых закономерных процессов, протекающих в них. Она есть номогенез, развитие по твердым законам».
По мнению Л. С. Берга, основными причинами эволюции являются изменения стереохимических свойств белков протоплазмы живых существ. Эти изменения приводят к развитию в определенном направлении.
Л. С. Берг писал, «что процесс эволюции идет закономерно, что он покоится на основе номогенеза. Как проявляются эти закономерности, мы видим, но почему они таковы, — это пока скрыто от нас. Равным образом, почему организмы в общем прогрессируют в своей организации, мы не знаем». Однако Л. С. Берг утверждал, что при этом «природа работает исключительно при помощи законов физики и химии».
Свою теорию Л. С. Берг иллюстрирует многочисленными примерами, взятыми из самых различных областей морфологии, и эти примеры, конечно, во всех случаях оказываются подтверждением его номогенеза.
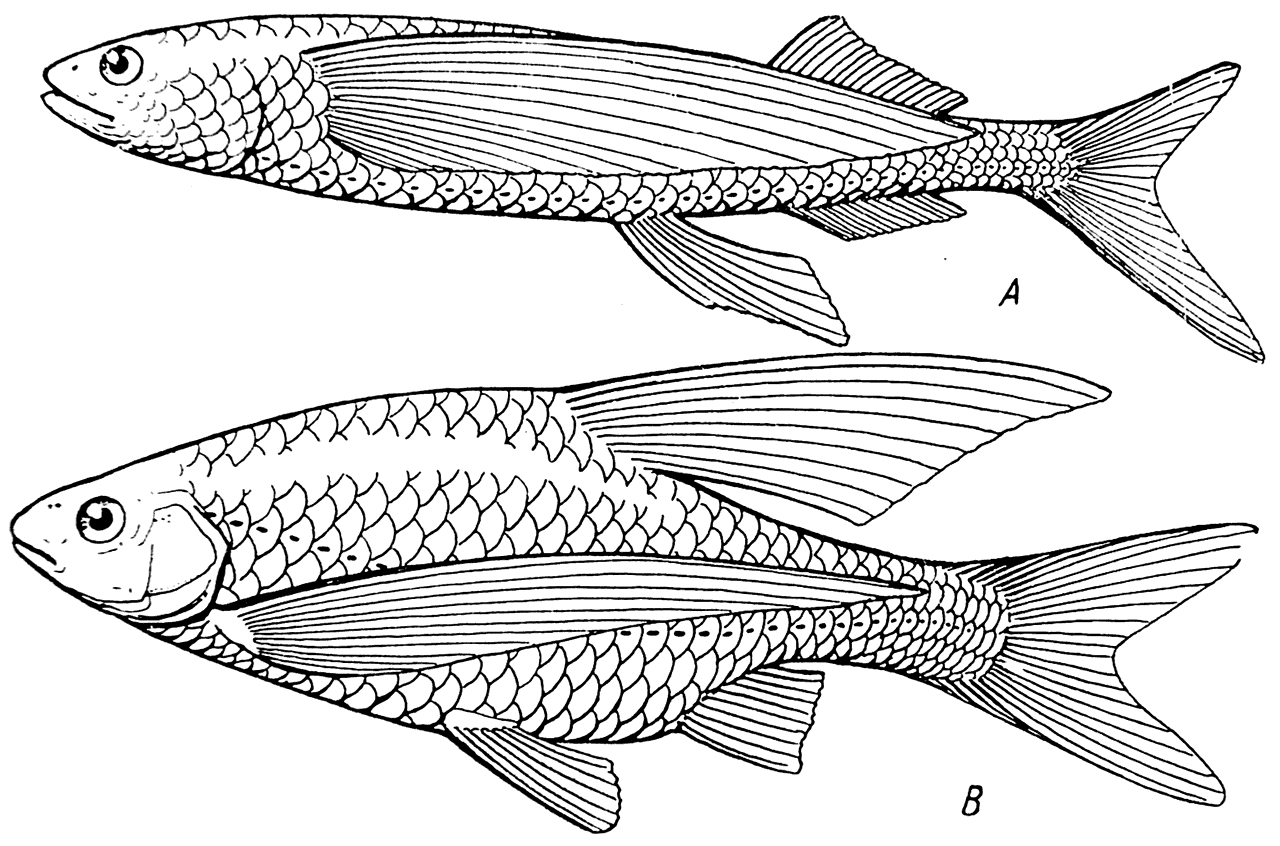
Рис. 14.
A — Exocoetus volitans; B — Rutilus rutilus aralensis monstrositas exocoetoides (по Л. С. Бергу).
В частности, ссылаясь на описанный им в 1916 г. интересный экземпляр аральской плотвы (Rutilus rutilus aralensis monstrositas exocoetoides, рис. 14, B) с необычайно длинными грудными плавниками, напоминающими соответствующие плавники летающей рыбы Exocoetus volitans (рис. 14, A), он приходит к заключению, что планирующий полет у летающих рыб развился «на основе закономерностей, вне всякого участия как естественного отбора, так и употребления и неупотребления органа». «Удлиненные плавники, — говорит он, — образуются потому, что они должны образоваться».
Таким образом, эволюционный процесс, по мнению Л. С. Берга, представляет собой непосредственный результат особых закономерных изменений в самих живых организмах. Эти изменения, независимо от окружающей среды, всегда ведут эволюцию по такому пути, который приводит к тому, что должно было быть.
Если Додерлейн в 1888 г. рассматривал vis inertiae только как необходимое дополнение к дарвиновским факторам эволюции, то Абель (O. Abel) в 1929 г. считал, что «сила инерции» является во всех случаях единственной причиной эволюционного процесса. По его мнению, причиной филогенетических изменений живых существ является так называемый «биологический закон инерции» (Biologisches Trägheitsgesetz). Этот закон некоторые биологи называют «законом Абеля».
Хотя Абель, как и Додерлейн, говорил об инерции, однако он давал совершенно иное обоснование своим взглядам.
Абель считал, что в настоящее время у нас нет никаких оснований сомневаться, что все процессы в живых организмах подчинены законам физики и химии, а все физические и химические процессы могут рассматриваться, как движение. Поэтому все без исключения функции живых организмов представляют собой явления не только физико-химические, но и механические. Но так как одним из основных законов механики следует считать закон инерции, то, по мнению О. Абеля, все функции животных и растений и их эволюционное развитие полностью подчинены этому закону. Поэтому однажды взятое направление эволюции затем на всех своих этапах в силу этого закона остается неизменным. Таким образом, по теории О. Абеля эволюционные изменения являются внутренним свойством живых организмов.
«В настоящее время мы знаем, — говорит Абель, — что всякий функционирующий организм, так же как его отдельные части и даже сама протоплазма, находится в состоянии движения, потому что функция есть деятельность и, следовательно, — движение. Но если это так, то все части организма должны подчиняться законам физики и, следовательно, механики, к которым в первую очередь относится закон инерции. Этот закон и для органического мира следует понимать в механическом, а не в переносном смысле».
Разумеется, мы не можем отрицать наличия физических и химических процессов в живых организмах. Но такие факторы эволюции как борьба за существование и естественный отбор не могут быть признаны ни физическими, ни химическими явлениями и они не встречаются в мертвой природе. А без этих факторов никакие физические и химические процессы, протекающие в живых организмах, не смогут вызвать их эволюционных изменений.
Таким образом, можно уверенно сказать, что физико-химические процессы в живых организмах также не приводят и не приводили к эволюционным изменениям, как они не превращали в эволюцию никакие изменения в неорганическом мире.
В 1933—1934 гг. Осборн (Н. F. Osborn) опубликовал свою теорию «аристогенеза» (ἄριστος — aristos — наилучший; γένεσις — genesis — происхождение).
Изучая в течение многих лет титанотериев и ископаемых хоботных, Осборн на большом материале проследил основные этапы эволюции этих млекопитающих. При этом у наиболее древних форм титанотериев он наблюдал появление небольших костных выступов на переднем отделе черепа, которые у более поздних форм превратились в хорошо выраженные рога (рис. 15).
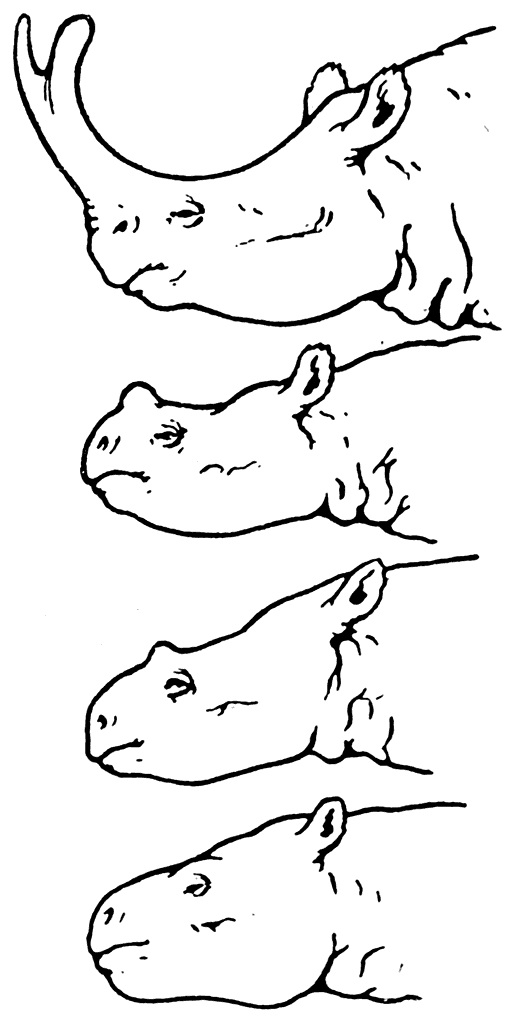
Рис. 15. Эволюционные изменения титанотериев.
У хоботных Осборн особенно детально изучал постепенное увеличение количества бугорков и гребней на жевательной поверхности зубов.
Постепенное увеличение размеров рогов у титанотериев и постепенное усложнение строения зубов у хоботных Осборн считал результатом особых изменений в плазме половых клеток этих животных. Эти изменения, по его мнению, выражались в появлении так называемых «аристогенов», которые затем медленно развивались и были причиной возникновения у животных различных полезных и притом постепенно улучшающихся особенностей.
«Мы знаем, — говорит Осборн, — как появляются эти аристогены, но мы не знаем, почему они появляются, т. е. причины их возникновения». «То, что вызывает аристоген в генеплазме, — пишет он, — для нас также таинственно, как и то, что вызывает зачаток рога на черепе титанотерия».
Однако, несмотря на всю эту таинственность, Осборн признавал, что «аристогены, конечно, не появляются самопроизвольно или без какой-либо прежде существующей или побуждающей причины». Они, по его мнению, возникают в результате способности животного реагировать приспособительно на различные особенности внешней среды и на борьбу с другими животными. В результате этого изменение пищи вызвало у хоботных появление аристогенов, обусловливающих постепенное усложнение зубов, а борьба с другими животными привела у титанотериев к возникновению аристогенов, которые были причиной появления и постепенного увеличения рогов.
Несмотря на то, что появление аристогенов в генеплазме животных, по мнению Осборна, как-то связано с внешней средой, сам эволюционный процесс (aristogenesis) все же, по его убеждению, является только результатом развития аристогенов.
«Аристогенез,— говорит он, — есть творческий процесс, вызванный совершенно новым биохимизмом зародышевой плазмы, процесс направленный, постепенный, прямой, идущий по пути к будущему приспособлению».
Таким образом, по теории Осборна, всякие эволюционные изменения предрешены биохимическими реакциями, происходящими в плазме половых клеток живых существ.
Все авторы, предлагающие ту или иную теорию эволюции для замены неудовлетворяющего их дарвинизма, в подтверждение своих взглядов, разумеется, всегда приводят некоторое количество фактических данных.
По моему мнению, нет ничего удивительного в том, что это оказывается возможным. Строение как современных, так и ископаемых животных настолько разнообразно, что если не пытаться выяснить в каждом отдельном случае ту конкретную обстановку, в которой формировались все морфологические особенности данного организма, то при желании можно без особого труда найти неограниченное количество доказательств в подтверждение правильности любой антидарвиновской гипотезы. Мало того, наши знания конкретной истории формирования многих анатомических особенностей животных настолько недостаточны, что одну и ту же морфологическую структуру можно легко трактовать с различных точек зрения.
Это можно показать хотя бы на следующем примере.
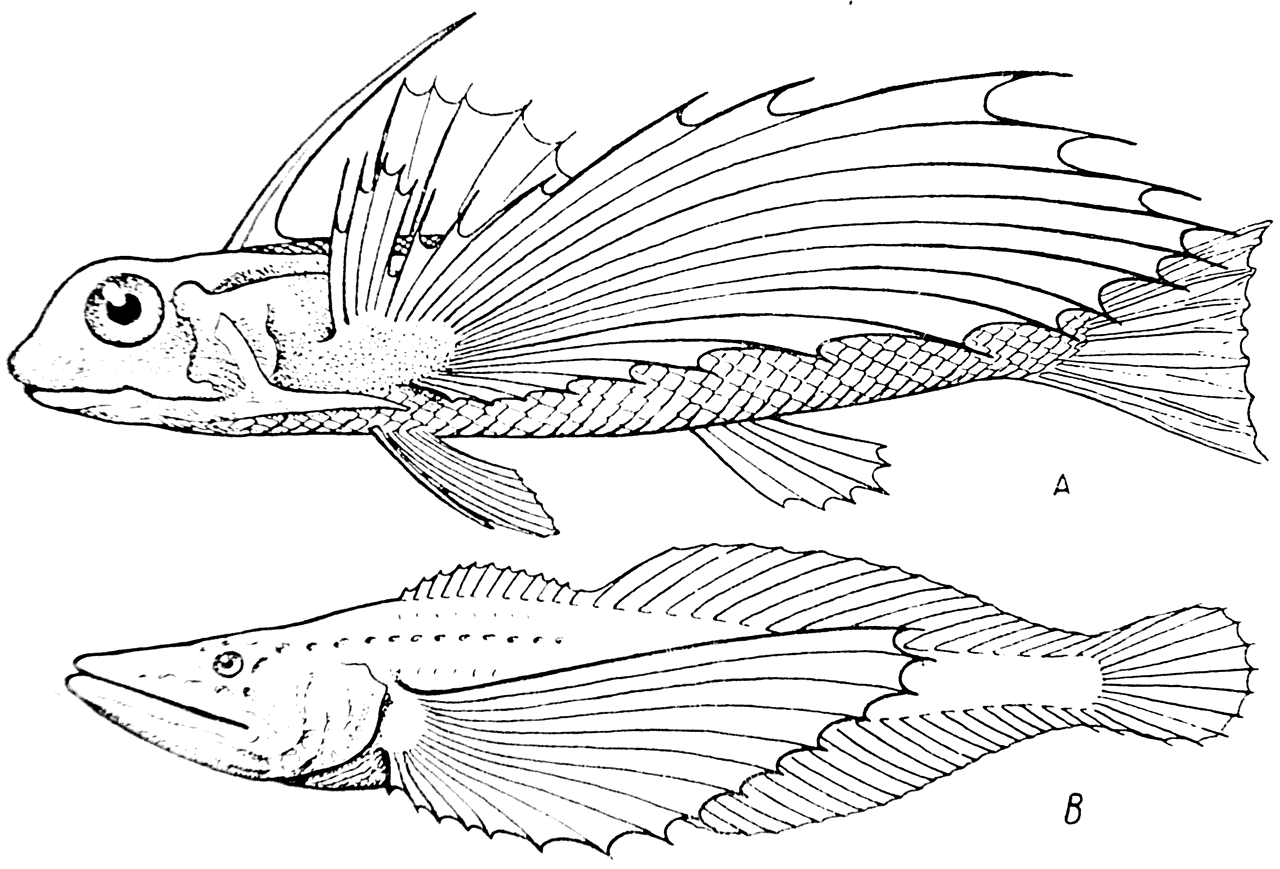
Рис. 16.
A — Dactylopterus orietitalis; B — Comephorus dybowskii.
В свое время в озере Байкал была обнаружена своеобразная рыба Comephorus dybowskii (рис. 16, В), у которой имелись очень большие грудные плавники, напоминающие соответствующие плавники летающей рыбы Dactylopterus orientalis (рис. 16, A). Причина образования и назначение, т. е. функция, этих плавников пока еще совершенно не выяснены. Поэтому ничто не мешает нам утверждать, что под влиянием внешней среды в плазме этой рыбы в свое время произошли такие изменения, которые привели к увеличению грудных плавников, которые, однако, аппаратом для планирующего полета пока еще не служат. Но дальнейшее их развитие, обусловленное дальнейшим органическим ростом плазмы, надо думать, будет идти в том же направлении, а это в конце концов неминуемо должно будет привести к появлению нового вида летающих рыб. Таким образом, очевидно, что Comephorus подтверждает ортогенез Эймера.
Однако в данном случае мы можем, соглашаясь с мнением Эймера, что развитие грудных плавников у Comephorus обусловлено физико-химическими изменениями плазмы рыбы, в то же время утверждать, что эти изменения наступили совершенно независимо от каких бы то ни было особенностей внешней среды. Мы можем предполагать, что они были обусловлены закономерными процессами, постоянно протекающими в плазме живых существ. Поэтому длинные плавники образовались только потому, что они должны были образоваться. Следовательно, байкальский Comephorus dybowskii является хорошей иллюстрацией не ортогенеза Т. Эймера, а номогенеза Л. С. Берга.
Впрочем, если принять во внимание, что все физико-химические процессы сопровождаются движением, а основным законом движения является закон инерции, то никто не может лишить нас права считать, что эволюционный процесс направляется именно этим законом и что развитие плавников у Comephorus шло по существу под влиянием только силы инерции. Эта сила и обусловливала и направляла эволюцию рыбы по строго определенному пути, что, в частности, привело к образованию у нее больших грудных плавников, а в будущем даст ей возможность превратиться в летающую рыбу…
Если это действительно так, то Comephorus dybowskii представляет собой прекрасный пример, подтверждающий не номогенез Л. С. Берга, а биологический закон инерции О. Абеля.
Наконец, у нас имеются все основания ссылаться на Comephorus как на очевидное доказательство того, что в генеплазме живых существ возникают аристогены, обусловливающие эволюцию. В данном случае мы имеем дело с аристогеном, вызвавшим появление больших грудных плавников. Дальнейшая эволюция этого аристогена неминуемо должна будет привести к возникновению байкальской летающей рыбы, вполне приспособленной ко всем особенностям жизни в этом озере. Поэтому мы можем считать, что Comephorus dybowskii является подтверждением не закона Абеля, а аристогенеза Осборна.
Все это можно утверждать, но ничего этого нельзя доказать. При этом следует иметь в виду, что интересные морфологические особенности Comephorus dybowskii, с одинаковой легкостью подтверждающие различные антидарвиновские гипотезы, в то же время не могут служить доказательством правильности только одной теории, именно: теории Ч. Дарвина, так как для этого совершенно необходимо всестороннее и детальное знание той конкретной и, несомненно, очень сложной обстановки, в которой на протяжении не одного миллиона лет совершалась эволюция этой рыбы.
Берг Л. С. Номогенез или эволюция на основе закономерностей. Труды Географического института, т. 1, 1922.
Ламарк Ж. Б. Философия зоологии. 1809. М.— Л., 1933.
Abel О. Palaeobiologie und Stammesgeschichte. Iena, 1929.
Dоdeгlein L. Phylogenetische Betrachtungen. Biologisches Centralblatt, Bd. 7, 1888.
Eimeг Th. Orthogenesis der Schmetterlinge. Ein Beweis bestimmt gerichteter Entwicklung und Ohnmacht der naturlichen Zuchtwahl bei der Artbildung. Leipzig, 1897.
Оsbоrn H. F. Aristogenesis, the creative principle in the origin of species. The American Naturalist, vol. 68, N 716, 1934.
Riggs E. S. Preliminary description of a new marsupial sabertooth from the pliocene of Argentina. Geological Series of Field Museum of Natural History, vol. 6, 1933.
Scott W. B. and Jepsen G. L. The mammalian fauna of the White River Oligocene. Part I. Insectivora and Carnivora. Transactions of the American Philosophical Society, vol. 28, 1936.